

The glycolytic enzyme phosphofructokinase-1 assembles into filaments
- PMID: 28646105
- PMCID: PMC5551713
- DOI: 10.1083/jcb.201701084
The glycolytic enzyme phosphofructokinase-1 assembles into filaments
Abstract
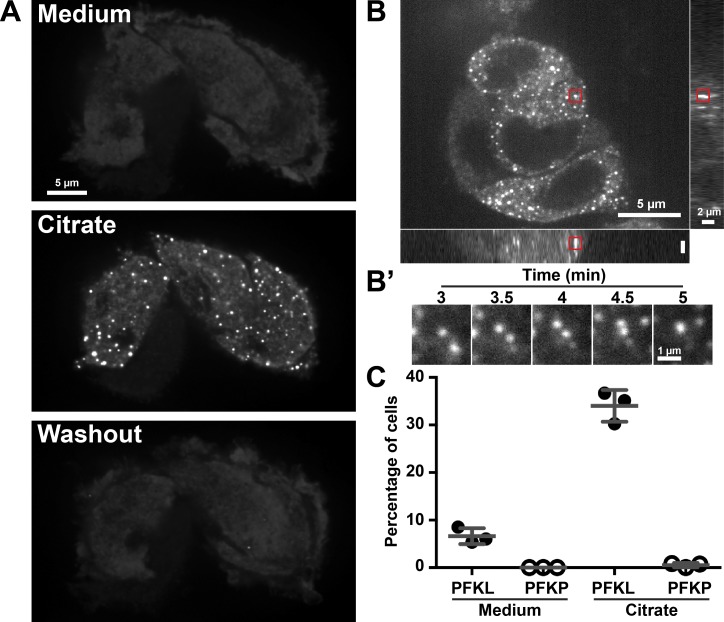
Despite abundant knowledge of the regulation and biochemistry of glycolytic enzymes, we have limited understanding on how they are spatially organized in the cell. Emerging evidence indicates that nonglycolytic metabolic enzymes regulating diverse pathways can assemble into polymers. We now show tetramer- and substrate-dependent filament assembly by phosphofructokinase-1 (PFK1), which is considered the "gatekeeper" of glycolysis because it catalyzes the step committing glucose to breakdown. Recombinant liver PFK1 (PFKL) isoform, but not platelet PFK1 (PFKP) or muscle PFK1 (PFKM) isoforms, assembles into filaments. Negative-stain electron micrographs reveal that filaments are apolar and made of stacked tetramers oriented with exposed catalytic sites positioned along the edge of the polymer. Electron micrographs and biochemical data with a PFKL/PFKP chimera indicate that the PFKL regulatory domain mediates filament assembly. Quantified live-cell imaging shows dynamic properties of localized PFKL puncta that are enriched at the plasma membrane. These findings reveal a new behavior of a key glycolytic enzyme with insights on spatial organization and isoform-specific glucose metabolism in cells.
© 2017 Webb et al.
Figures
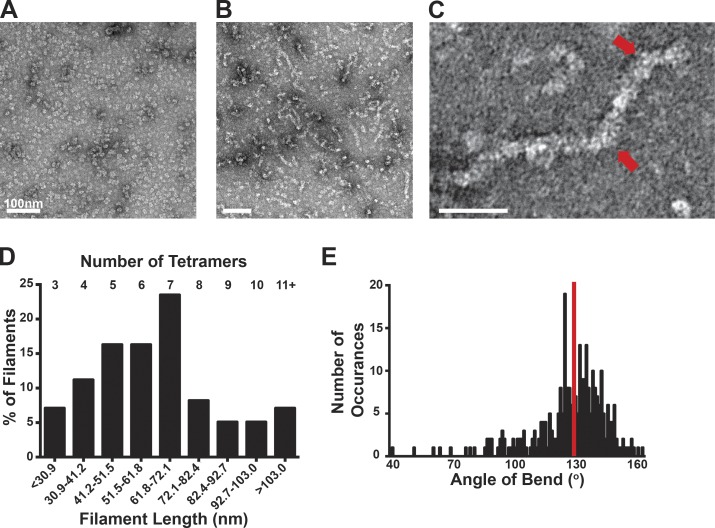
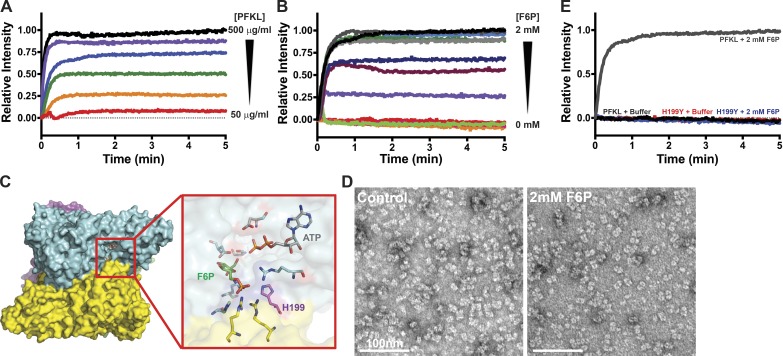
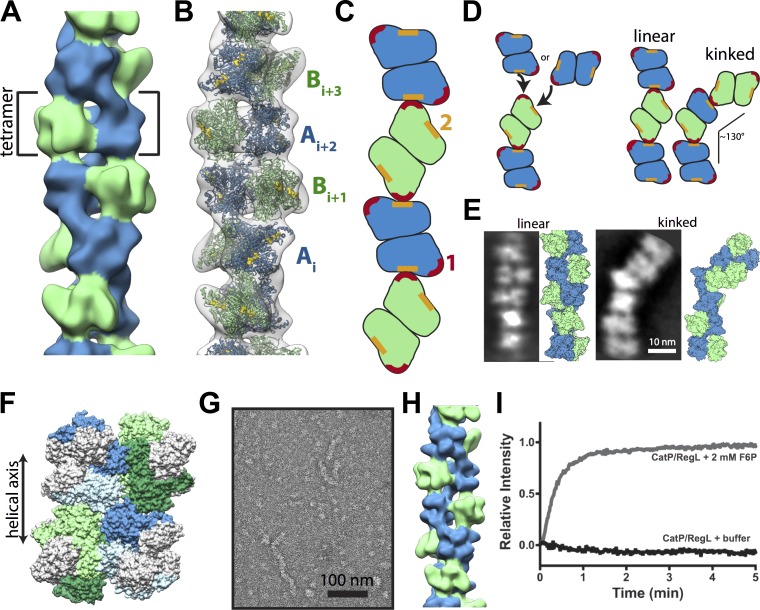
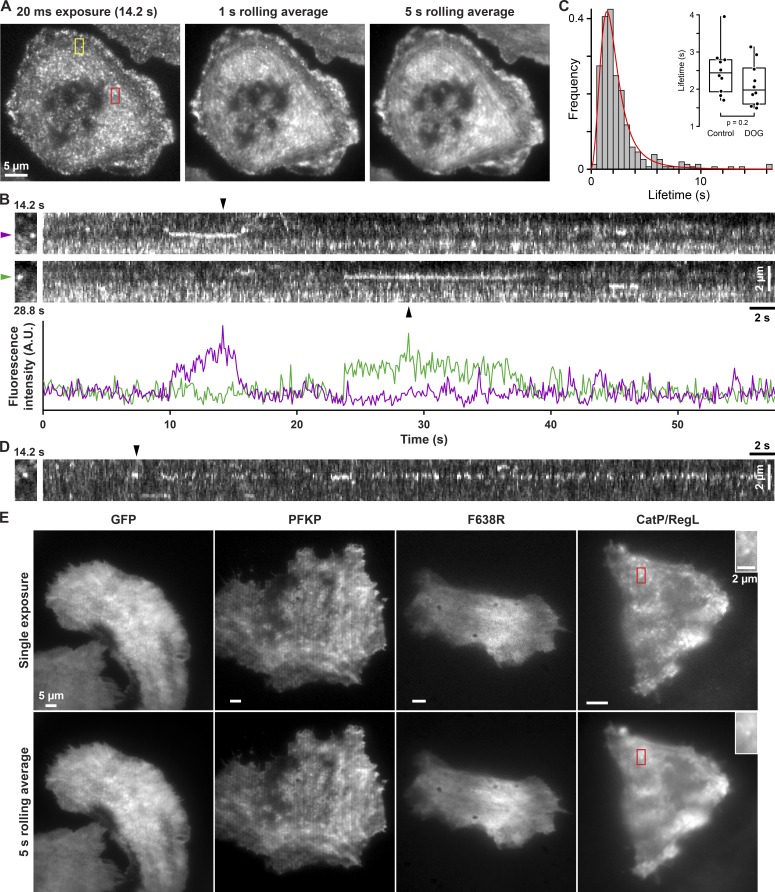
Comment in
-
Strength in numbers: Phosphofructokinase polymerization prevails in the liver.J Cell Biol. 2017 Aug 7;216(8):2239-2241. doi: 10.1083/jcb.201706005. Epub 2017 Jul 17. J Cell Biol. 2017. PMID: 28716843 Free PMC article.
References
-
- Beaty N.B., and Lane M.D.. 1983. The polymerization of acetyl-CoA carboxylase. J. Biol. Chem. 258:13051–13055. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
