

Modulation of oxidative phosphorylation and redox homeostasis in mitochondrial NDUFS4 deficiency via mesenchymal stem cells
- PMID: 28646906
- PMCID: PMC5482938
- DOI: 10.1186/s13287-017-0601-7
Modulation of oxidative phosphorylation and redox homeostasis in mitochondrial NDUFS4 deficiency via mesenchymal stem cells
Abstract
Background: Disorders of the oxidative phosphorylation (OXPHOS) system represent a large group among the inborn errors of metabolism. The most frequently observed biochemical defect is isolated deficiency of mitochondrial complex I (CI). No effective treatment strategies for CI deficiency are so far available. The purpose of this study was to investigate whether and how mesenchymal stem cells (MSCs) are able to modulate metabolic function in fibroblast cell models of CI deficiency.
Methods: We used human and murine fibroblasts with a defect in the nuclear DNA encoded NDUFS4 subunit of CI. Fibroblasts were co-cultured with MSCs under different stress conditions and intercellular mitochondrial transfer was assessed by flow cytometry and fluorescence microscopy. Reactive oxygen species (ROS) levels were measured using MitoSOX-Red. Protein levels of CI were analysed by blue native polyacrylamide gel electrophoresis (BN-PAGE).
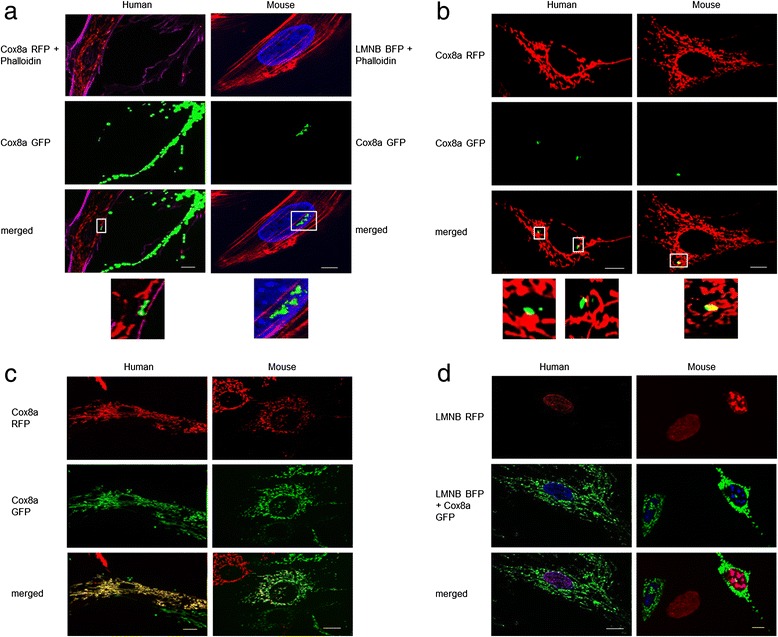
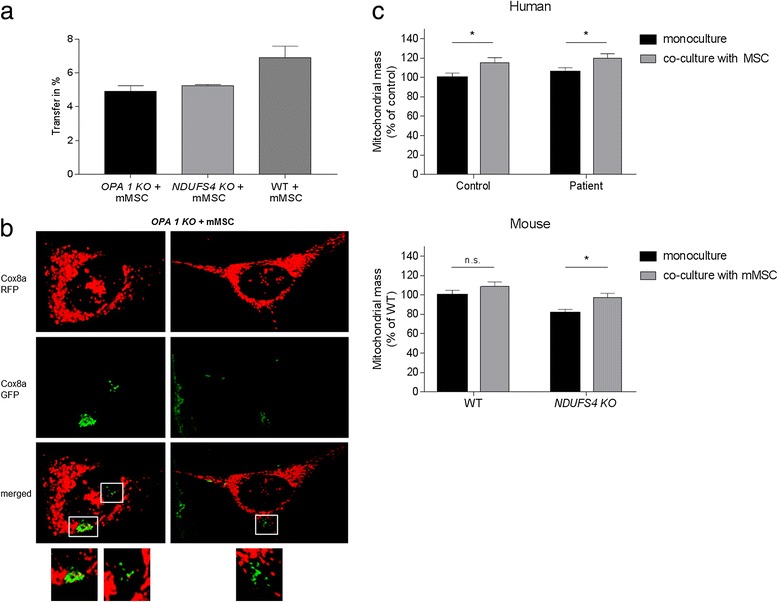
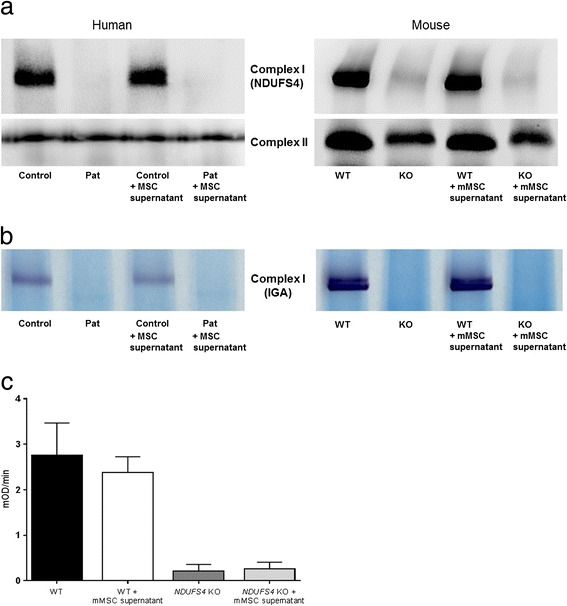
Results: Direct cellular interactions and mitochondrial transfer between MSCs and human as well as mouse fibroblast cell lines were demonstrated. Mitochondrial transfer was visible in 13.2% and 6% of fibroblasts (e.g. fibroblasts containing MSC mitochondria) for human and mouse cell lines, respectively. The transfer rate could be further stimulated via treatment of cells with TNF-α. MSCs effectively lowered cellular ROS production in NDUFS4-deficient fibroblast cell lines (either directly via co-culture or indirectly via incubation of cell lines with cell-free MSC supernatant). However, CI protein expression and activity were not rescued by MSC treatment.
Conclusion: This study demonstrates the interplay between MSCs and fibroblast cell models of isolated CI deficiency including transfer of mitochondria as well as modulation of cellular ROS levels. Further exploration of these cellular interactions might help to develop MSC-based treatment strategies for human CI deficiency.
Figures


References
-
- Distelmaier F, Valsecchi F, Liemburg-Apers DC, Lebiedzinska M, Rodenburg RJ, Heil S, Keijer J, Fransen J, Imamura H, Danhauser K, et al. Mitochondrial dysfunction in primary human fibroblasts triggers an adaptive cell survival program that requires AMPK-alpha. Biochim Biophys Acta. 2015;1852(3):529–40. doi: 10.1016/j.bbadis.2014.12.012. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
