

Dynamic Reorganization of Chromatin Accessibility Signatures during Dedifferentiation of Secretory Precursors into Lgr5+ Intestinal Stem Cells
- PMID: 28648363
- PMCID: PMC5505276
- DOI: 10.1016/j.stem.2017.05.001
Dynamic Reorganization of Chromatin Accessibility Signatures during Dedifferentiation of Secretory Precursors into Lgr5+ Intestinal Stem Cells
Abstract
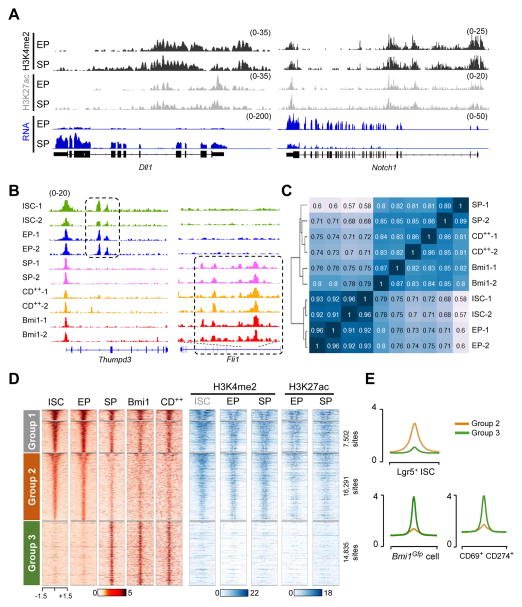
Replicating Lgr5+ stem cells and quiescent Bmi1+ cells behave as intestinal stem cells (ISCs) in vivo. Disrupting Lgr5+ ISCs triggers epithelial renewal from Bmi1+ cells, from secretory or absorptive progenitors, and from Paneth cell precursors, revealing a high degree of plasticity within intestinal crypts. Here, we show that GFP+ cells from Bmi1GFP mice are preterminal enteroendocrine cells and we identify CD69+CD274+ cells as related goblet cell precursors. Upon loss of native Lgr5+ ISCs, both populations revert toward an Lgr5+ cell identity. While active histone marks are distributed similarly between Lgr5+ ISCs and progenitors of both major lineages, thousands of cis elements that control expression of lineage-restricted genes are selectively open in secretory cells. This accessibility signature dynamically converts to that of Lgr5+ ISCs during crypt regeneration. Beyond establishing the nature of Bmi1GFP+ cells, these findings reveal how chromatin status underlies intestinal cell diversity and dedifferentiation to restore ISC function and intestinal homeostasis.
Keywords: accessible chromatin; cell plasticity; chromatin modulation; dedifferentiation; facultative stem cells; intestinal stem cells; reserve stem cells.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures





References
-
- Barker N. Adult intestinal stem cells: critical drivers of epithelial homeostasis and regeneration. Nat Rev Mol Cell Biol. 2014;15:19–33. - PubMed
-
- Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–1007. - PubMed
-
- Buczacki SJ, Zecchini HI, Nicholson AM, Russell R, Vermeulen L, Kemp R, Winton DJ. Intestinal label-retaining cells are secretory precursors expressing Lgr5. Nature. 2013;495:65–69. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
