

A tripartite complex of suPAR, APOL1 risk variants and αvβ3 integrin on podocytes mediates chronic kidney disease
- PMID: 28650456
- PMCID: PMC6019326
- DOI: 10.1038/nm.4362
A tripartite complex of suPAR, APOL1 risk variants and αvβ3 integrin on podocytes mediates chronic kidney disease
Abstract
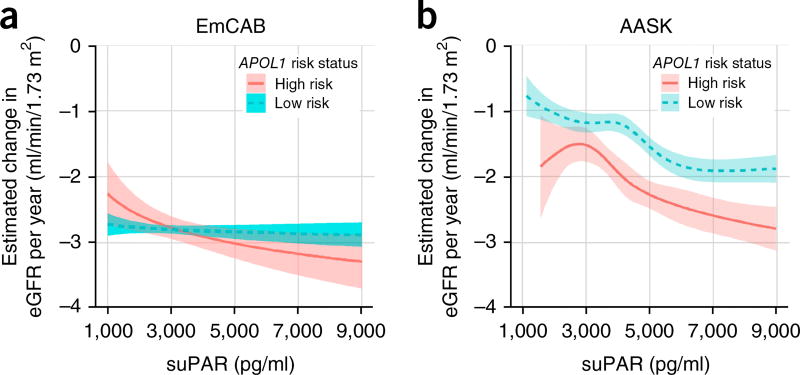
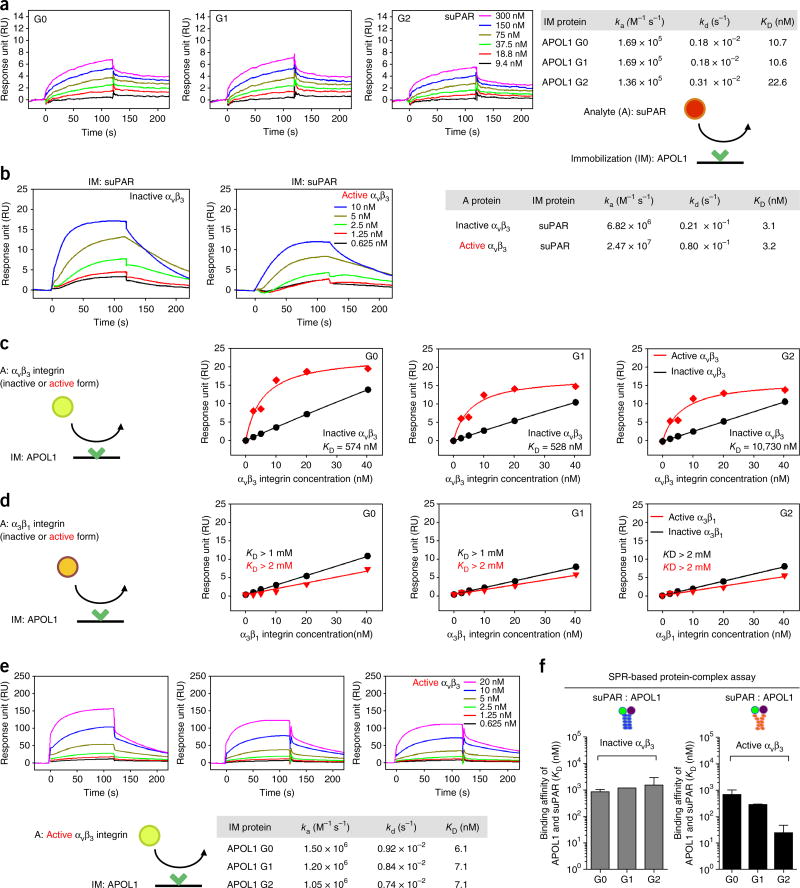
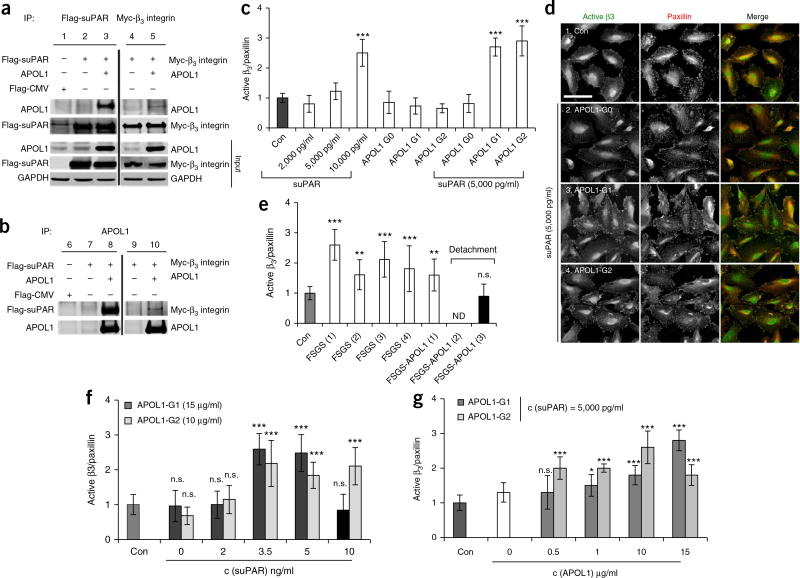
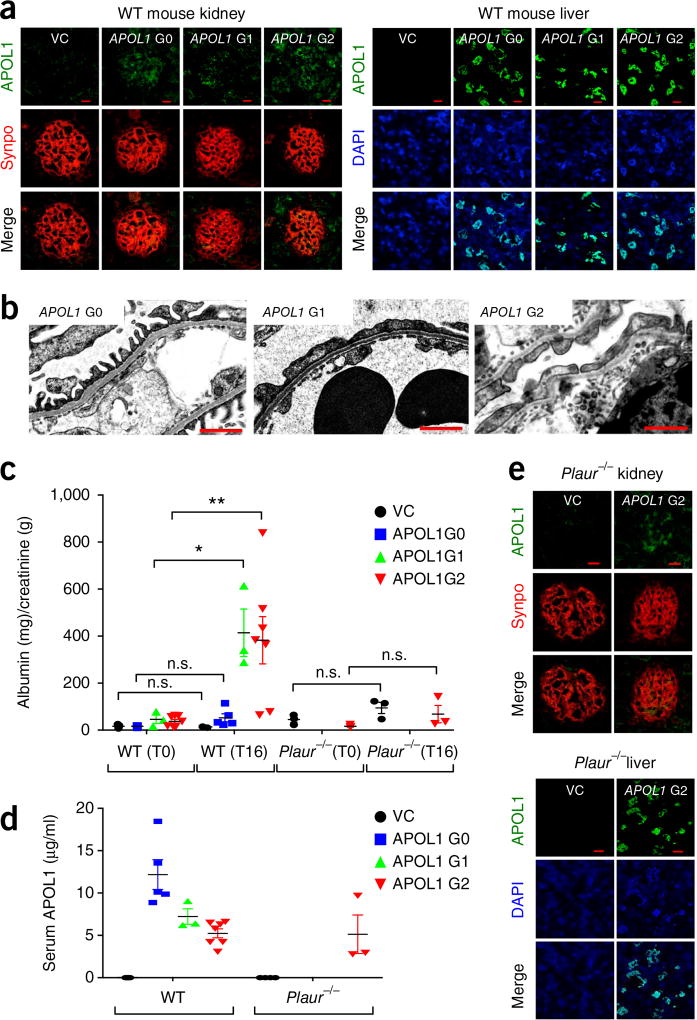
Soluble urokinase plasminogen activator receptor (suPAR) independently predicts chronic kidney disease (CKD) incidence and progression. Apolipoprotein L1 (APOL1) gene variants G1 and G2, but not the reference allele (G0), are associated with an increased risk of CKD in individuals of recent African ancestry. Here we show in two large, unrelated cohorts that decline in kidney function associated with APOL1 risk variants was dependent on plasma suPAR levels: APOL1-related risk was attenuated in patients with lower suPAR, and strengthened in those with higher suPAR levels. Mechanistically, surface plasmon resonance studies identified high-affinity interactions between suPAR, APOL1 and αvβ3 integrin, whereby APOL1 protein variants G1 and G2 exhibited higher affinity for suPAR-activated avb3 integrin than APOL1 G0. APOL1 G1 or G2 augments αvβ3 integrin activation and causes proteinuria in mice in a suPAR-dependent manner. The synergy of circulating factor suPAR and APOL1 G1 or G2 on αvβ3 integrin activation is a mechanism for CKD.
Conflict of interest statement
The authors declare competing financial interests: details are available in the
Figures
Comment in
-
Chronic kidney disease: Role of suPAR in APOL1-associated kidney disease.Nat Rev Nephrol. 2017 Sep;13(9):514. doi: 10.1038/nrneph.2017.97. Epub 2017 Jul 3. Nat Rev Nephrol. 2017. PMID: 28669990 No abstract available.
References
-
- Chen TK, Estrella MM, Parekh RS. The evolving science of apolipoprotein-L1 and kidney disease. Curr. Opin. Nephrol. Hypertens. 2016;25:217–225. - PubMed
MeSH terms
Substances
Grants and funding
- R56 HL126558/HL/NHLBI NIH HHS/United States
- R01 DK106051/DK/NIDDK NIH HHS/United States
- R01 HL089650/HL/NHLBI NIH HHS/United States
- R01 DK101350/DK/NIDDK NIH HHS/United States
- R01 NS064162/NS/NINDS NIH HHS/United States
- P01 HL086773/HL/NHLBI NIH HHS/United States
- DP3 DK094346/DK/NIDDK NIH HHS/United States
- U01 DK085689/DK/NIDDK NIH HHS/United States
- R01 DK108803/DK/NIDDK NIH HHS/United States
- R01 HL095479/HL/NHLBI NIH HHS/United States
- RF1 AG051633/AG/NIA NIH HHS/United States
- UL1 RR025008/RR/NCRR NIH HHS/United States
- R01 DK087961/DK/NIDDK NIH HHS/United States
- U10 HL110302/HL/NHLBI NIH HHS/United States
- P20 HL113451/HL/NHLBI NIH HHS/United States
- P01 HL101398/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
