

A New Facet of Vitamin B12: Gene Regulation by Cobalamin-Based Photoreceptors
- PMID: 28654327
- PMCID: PMC7153952
- DOI: 10.1146/annurev-biochem-061516-044500
A New Facet of Vitamin B12: Gene Regulation by Cobalamin-Based Photoreceptors
Abstract
Living organisms sense and respond to light, a crucial environmental factor, using photoreceptors, which rely on bound chromophores such as retinal, flavins, or linear tetrapyrroles for light sensing. The discovery of photoreceptors that sense light using 5'-deoxyadenosylcobalamin, a form of vitamin B12 that is best known as an enzyme cofactor, has expanded the number of known photoreceptor families and unveiled a new biological role of this vitamin. The prototype of these B12-dependent photoreceptors, the transcriptional repressor CarH, is widespread in bacteria and mediates light-dependent gene regulation in a photoprotective cellular response. CarH activity as a transcription factor relies on the modulation of its oligomeric state by 5'-deoxyadenosylcobalamin and light. This review surveys current knowledge about these B12-dependent photoreceptors, their distribution and mode of action, and the structural and photochemical basis of how they orchestrate signal transduction and control gene expression.
Keywords: CarH; chromophore; optogenetics; photochemistry; photoregulation; transcriptional repressor.
Figures

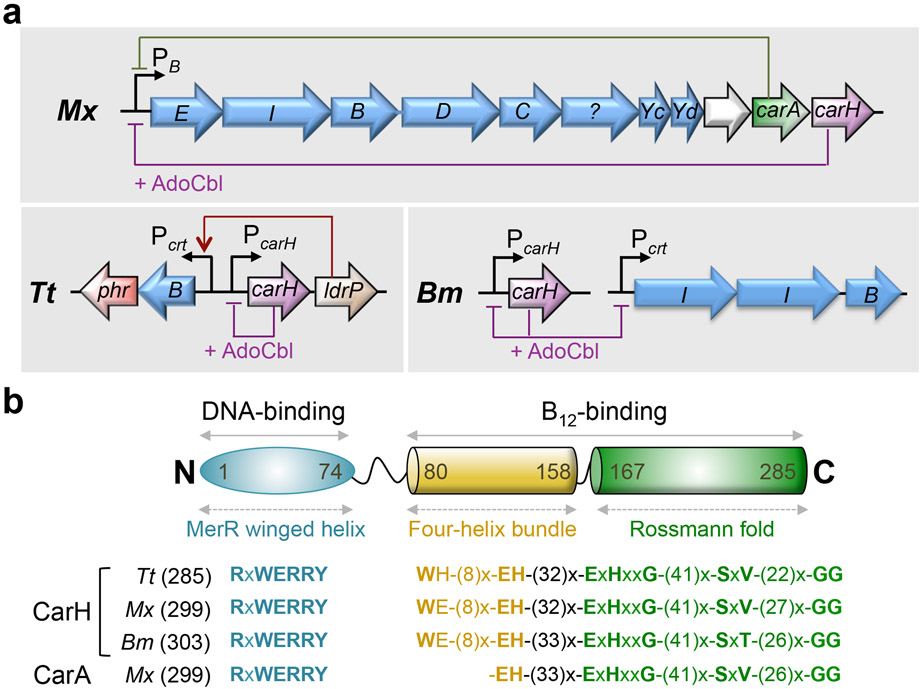


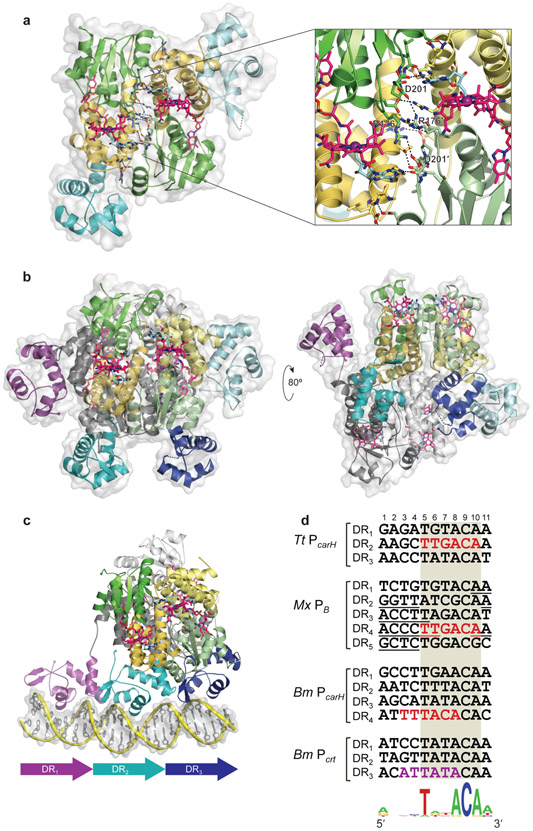


Similar articles
-
Plasticity in oligomerization, operator architecture, and DNA binding in the mode of action of a bacterial B12-based photoreceptor.J Biol Chem. 2018 Nov 16;293(46):17888-17905. doi: 10.1074/jbc.RA118.004838. Epub 2018 Sep 27. J Biol Chem. 2018. PMID: 30262667 Free PMC article.
-
Vitamin B12 photoreceptors.Vitam Horm. 2022;119:149-184. doi: 10.1016/bs.vh.2022.01.007. Epub 2022 Mar 10. Vitam Horm. 2022. PMID: 35337618 Review.
-
Light-dependent gene regulation by a coenzyme B12-based photoreceptor.Proc Natl Acad Sci U S A. 2011 May 3;108(18):7565-70. doi: 10.1073/pnas.1018972108. Epub 2011 Apr 18. Proc Natl Acad Sci U S A. 2011. PMID: 21502508 Free PMC article.
-
Structural basis for gene regulation by a B12-dependent photoreceptor.Nature. 2015 Oct 22;526(7574):536-41. doi: 10.1038/nature14950. Epub 2015 Sep 28. Nature. 2015. PMID: 26416754 Free PMC article.
-
B12-based photoreceptors: from structure and function to applications in optogenetics and synthetic biology.Curr Opin Struct Biol. 2019 Aug;57:47-55. doi: 10.1016/j.sbi.2019.01.020. Epub 2019 Mar 7. Curr Opin Struct Biol. 2019. PMID: 30851578 Review.
Cited by
-
Photoglobin, a distinct family of non-heme binding globins, defines a potential photosensor in prokaryotic signal transduction systems.Comput Struct Biotechnol J. 2021 Dec 18;20:261-273. doi: 10.1016/j.csbj.2021.12.022. eCollection 2022. Comput Struct Biotechnol J. 2021. PMID: 35024098 Free PMC article.
-
Role and Function of Class III LitR, a Photosensor Homolog from Burkholderia multivorans.J Bacteriol. 2018 Nov 26;200(24):e00285-18. doi: 10.1128/JB.00285-18. Print 2018 Dec 15. J Bacteriol. 2018. PMID: 30249707 Free PMC article.
-
Light Modulates the Physiology of Nonphototrophic Actinobacteria.J Bacteriol. 2019 Apr 24;201(10):e00740-18. doi: 10.1128/JB.00740-18. Print 2019 May 15. J Bacteriol. 2019. PMID: 30692175 Free PMC article.
-
Milestones in the development of Myxococcus xanthus as a model multicellular bacterium.J Bacteriol. 2025 Jul 24;207(7):e0007125. doi: 10.1128/jb.00071-25. Epub 2025 Jun 17. J Bacteriol. 2025. PMID: 40525847 Free PMC article. Review.
-
Regulation of Gene Expression Through Effector-dependent Conformational Switching by Cobalamin Riboswitches.J Mol Biol. 2022 Sep 30;434(18):167585. doi: 10.1016/j.jmb.2022.167585. Epub 2022 Apr 12. J Mol Biol. 2022. PMID: 35427633 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
