

Three distinct 3-methylcytidine (m3C) methyltransferases modify tRNA and mRNA in mice and humans
- PMID: 28655767
- PMCID: PMC5582859
- DOI: 10.1074/jbc.M117.798298
Three distinct 3-methylcytidine (m3C) methyltransferases modify tRNA and mRNA in mice and humans
Abstract
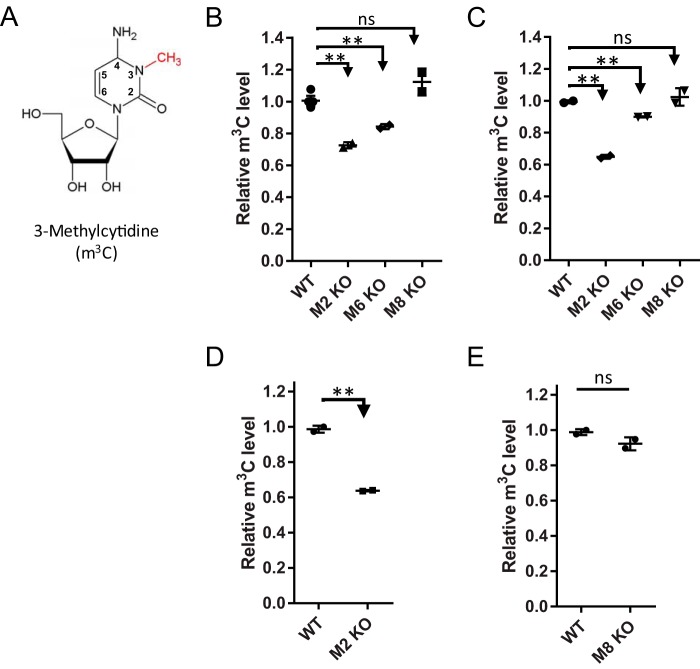
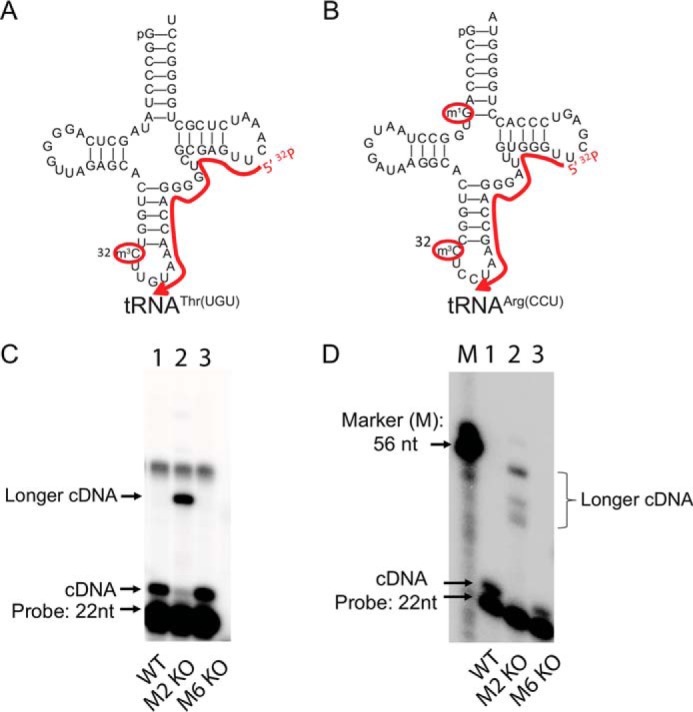
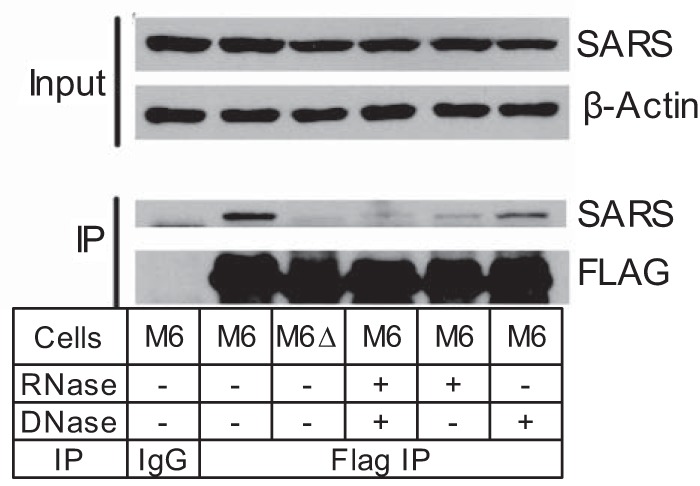
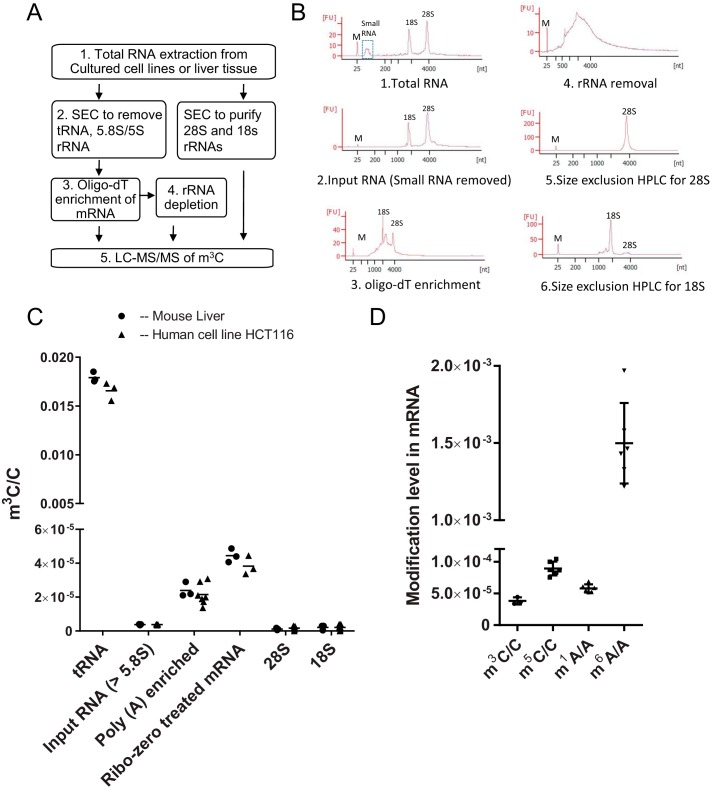
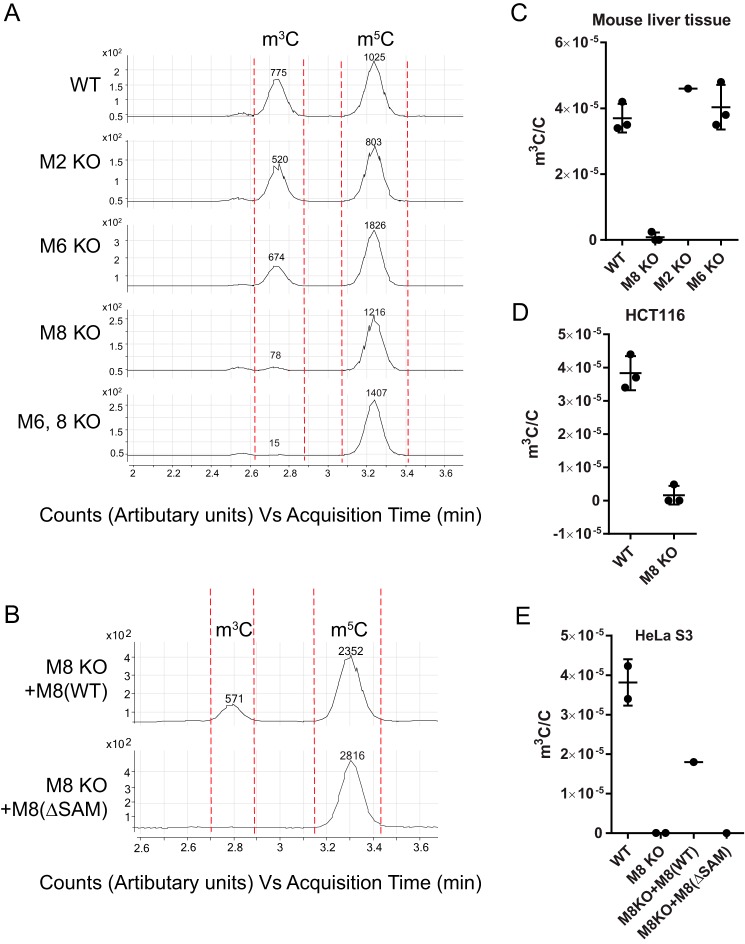
Chemical RNA modifications are central features of epitranscriptomics, highlighted by the discovery of modified ribonucleosides in mRNA and exemplified by the critical roles of RNA modifications in normal physiology and disease. Despite a resurgent interest in these modifications, the biochemistry of 3-methylcytidine (m3C) formation in mammalian RNAs is still poorly understood. However, the recent discovery of trm141 as the second gene responsible for m3C presence in RNA in fission yeast raises the possibility that multiple enzymes are involved in m3C formation in mammals as well. Here, we report the discovery and characterization of three distinct m3C-contributing enzymes in mice and humans. We found that methyltransferase-like (METTL) 2 and 6 contribute m3C in specific tRNAs and that METTL8 only contributes m3C to mRNA. MS analysis revealed that there is an ∼30-40% and ∼10-15% reduction, respectively, in METTL2 and -6 null-mutant cells, of m3C in total tRNA, and primer extension analysis located METTL2-modified m3C at position 32 of tRNAThr isoacceptors and tRNAArg(CCU) We also noted that METTL6 interacts with seryl-tRNA synthetase in an RNA-dependent manner, suggesting a role for METTL6 in modifying serine tRNA isoacceptors. METTL8, however, modified only mRNA, as determined by biochemical and genetic analyses in Mettl8 null-mutant mice and two human METTL8 mutant cell lines. Our findings provide the first evidence of the existence of m3C modification in mRNA, and the discovery of METTL8 as an mRNA m3C writer enzyme opens the door to future studies of other m3C epitranscriptomic reader and eraser functions.
Keywords: Mettl2; Mettl6; Mettl8; RNA methyltransferase; human; m3C; mRNA; mice; mouse; transfer RNA (tRNA).
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Comment in
-
A new modification for mammalian messenger RNA.J Biol Chem. 2017 Sep 1;292(35):14704-14705. doi: 10.1074/jbc.H117.798298. J Biol Chem. 2017. PMID: 28864714 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
