

Use of RNA-Protein Complexes for Genome Editing in Non- albicans Candida Species
- PMID: 28657070
- PMCID: PMC5480035
- DOI: 10.1128/mSphere.00218-17
Use of RNA-Protein Complexes for Genome Editing in Non- albicans Candida Species
Abstract
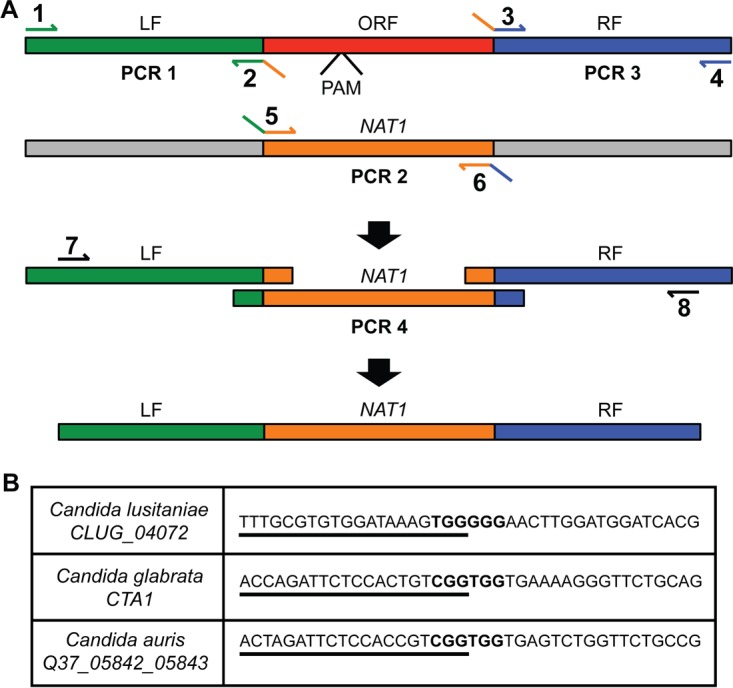
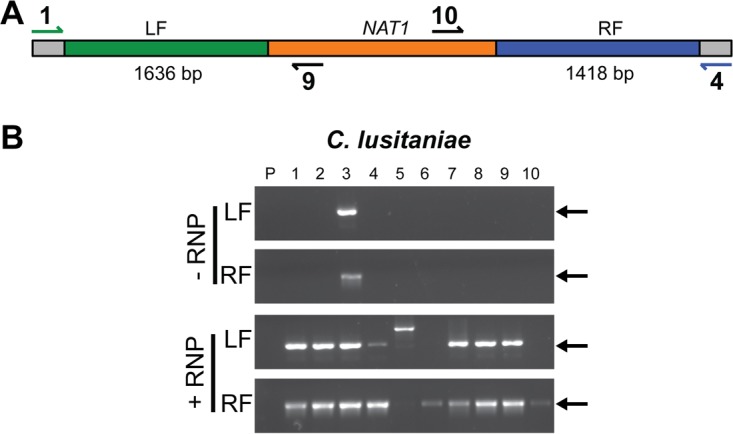
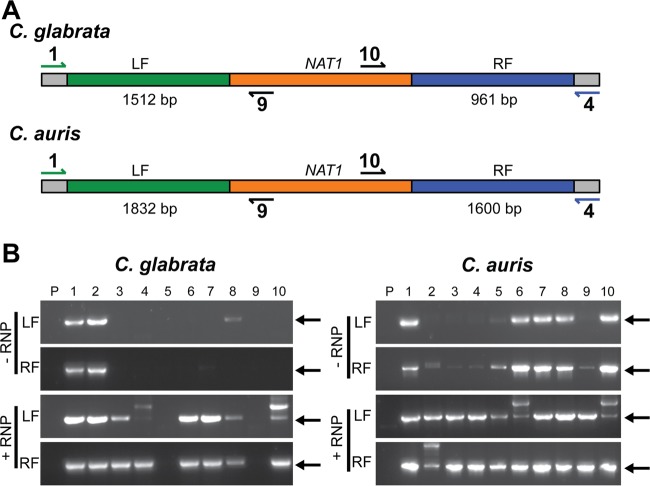
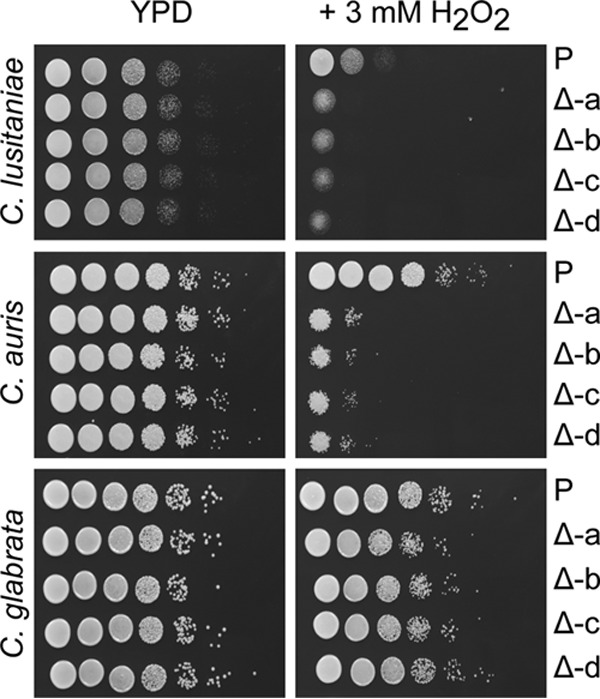
Clustered regularly interspaced short palindromic repeat (CRISPR)-Cas9 genome modification systems have greatly facilitated the genetic analysis of fungal pathogens. In CRISPR-Cas9 genome editing methods designed for use in Candida albicans, DNAs that encode the necessary components are expressed in the target cells. Unfortunately, expression constructs that work efficiently in C. albicans are not necessarily expressed well in other pathogenic species within the genus Candida or the related genus Clavispora. To circumvent the need for species-specific expression constructs, we implemented an expression-free CRISPR genome editing system and demonstrated its successful use in three different non-albicans Candida species: Candida (Clavispora) lusitaniae, Candida glabrata, and Candida auris. In CRISPR-Cas9-mediated genome editing methods, a targeted double-stranded DNA break can be repaired by homologous recombination to a template designed by the investigator. In this protocol, the DNA cleavage is induced upon transformation of purified Cas9 protein in complex with gene-specific and scaffold RNAs, referred to as RNA-protein complexes (RNPs). In all three species, the use of RNPs increased both the number of transformants and the percentage of transformants in which the target gene was successfully replaced with a selectable marker. We constructed mutants defective in known or putative catalase genes in C. lusitaniae, C. glabrata, and C. auris and demonstrated that, in all three species, mutants were more susceptible to hydrogen peroxide than the parental strain. This method, which circumvents the need for expression of CRISPR-Cas9 components, may be broadly useful in the study of diverse Candida species and emergent pathogens for which there are limited genetic tools. IMPORTANCE Existing CRISPR-Cas9 genome modification systems for use in Candida albicans, which rely on constructs to endogenously express the Cas9 protein and guide RNA, do not work efficiently in other Candida species due to inefficient promoter activity. Here, we present an expression-free method that uses RNA-protein complexes and demonstrate its use in three Candida species known for their drug resistance profiles. We propose that this system will aid the genetic analysis of fungi that lack established genetic systems.
Keywords: CRISPR; Candida; auris; genome editing; glabrata; lusitaniae; molecular methods.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
