

Development of a CRISPR-Cas9 System for Efficient Genome Editing of Candida lusitaniae
- PMID: 28657072
- PMCID: PMC5480034
- DOI: 10.1128/mSphere.00217-17
Development of a CRISPR-Cas9 System for Efficient Genome Editing of Candida lusitaniae
Abstract
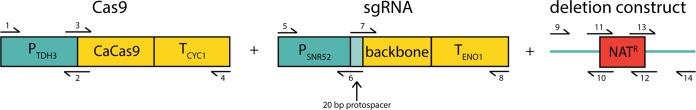
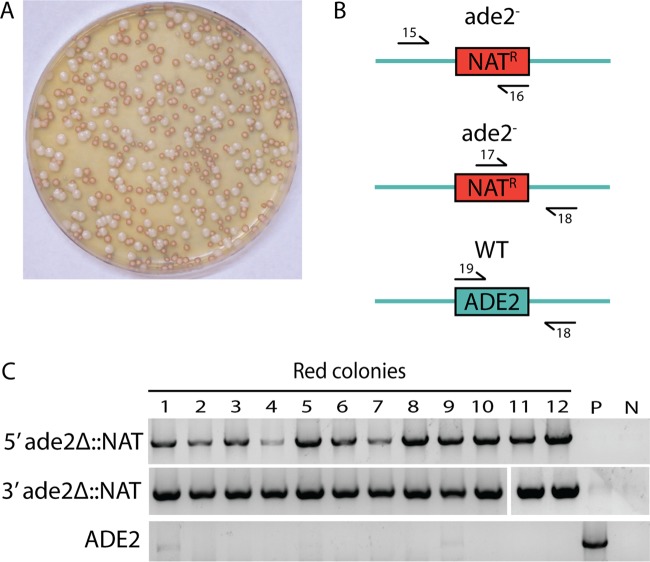
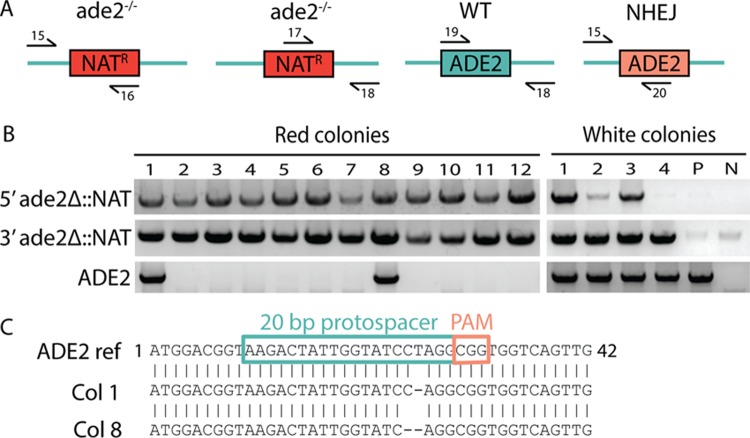
Candida lusitaniae is a member of the Candida clade that includes a diverse group of fungal species relevant to both human health and biotechnology. This species exhibits a full sexual cycle to undergo interconversion between haploid and diploid forms. C. lusitaniae is also an emerging opportunistic pathogen that can cause serious bloodstream infections in the clinic and yet has often proven to be refractory to facile genetic manipulations. In this work, we develop a clustered regularly interspaced short palindromic repeat (CRISPR) and CRISPR-associated gene 9 (Cas9) system to enable genome editing of C. lusitaniae. We demonstrate that expression of CRISPR-Cas9 components under species-specific promoters is necessary for efficient gene targeting and can be successfully applied to multiple genes in both haploid and diploid isolates. Gene deletion efficiencies with CRISPR-Cas9 were further enhanced in C. lusitaniae strains lacking the established nonhomologous end joining (NHEJ) factors Ku70 and DNA ligase 4. These results indicate that NHEJ plays an important role in directing the repair of DNA double-strand breaks (DSBs) in C. lusitaniae and that removal of this pathway increases integration of gene deletion templates by homologous recombination. The described approaches significantly enhance the ability to perform genetic studies in, and promote understanding of, this emerging human pathogen and model sexual species. IMPORTANCE The ability to perform efficient genome editing is a key development for detailed mechanistic studies of a species. Candida lusitaniae is an important member of the Candida clade and is relevant both as an emerging human pathogen and as a model for understanding mechanisms of sexual reproduction. We highlight the development of a CRISPR-Cas9 system for efficient genome manipulation in C. lusitaniae and demonstrate the importance of species-specific promoters for expression of CRISPR components. We also demonstrate that the NHEJ pathway contributes to non-template-mediated repair of DNA DSBs and that removal of this pathway enhances efficiencies of gene targeting by CRISPR-Cas9. These results therefore establish important genetic tools for further exploration of C. lusitaniae biology.
Keywords: CRISPR; DNA double-strand break; homology-directed repair; nonhomologous end joining.
Figures
References
-
- Butler G, Rasmussen MD, Lin MF, Santos MA, Sakthikumar S, Munro CA, Rheinbay E, Grabherr M, Forche A, Reedy JL, Agrafioti I, Arnaud MB, Bates S, Brown AJ, Brunke S, Costanzo MC, Fitzpatrick DA, de Groot PW, Harris D, Hoyer LL, Hube B, Klis FM, Kodira C, Lennard N, Logue ME, Martin R, Neiman AM, Nikolaou E, Quail MA, Quinn J, Santos MC, Schmitzberger FF, Sherlock G, Shah P, Silverstein KA, Skrzypek MS, Soll D, Staggs R, Stansfield I, Stumpf MP, Sudbery PE, Srikantha T, Zeng Q, Berman J, Berriman M, Heitman J, Gow NA, Lorenz MC, Birren BW, Kellis M. 2009. Evolution of pathogenicity and sexual reproduction in eight Candida genomes. Nature 459:657–662. doi: 10.1038/nature08064. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
