

Recurrent and functional regulatory mutations in breast cancer
- PMID: 28658208
- PMCID: PMC5563978
- DOI: 10.1038/nature22992
Recurrent and functional regulatory mutations in breast cancer
Abstract
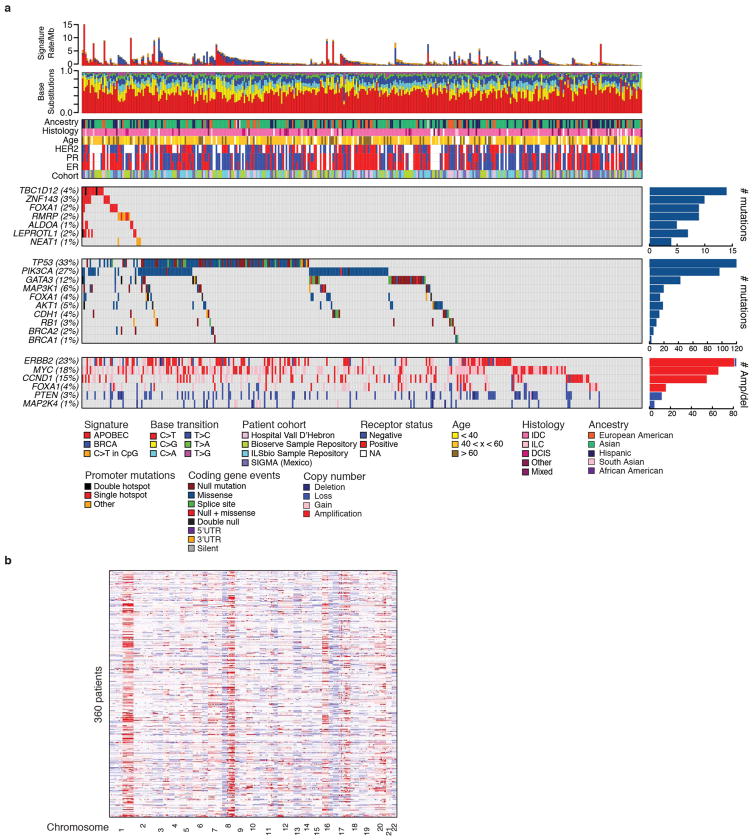
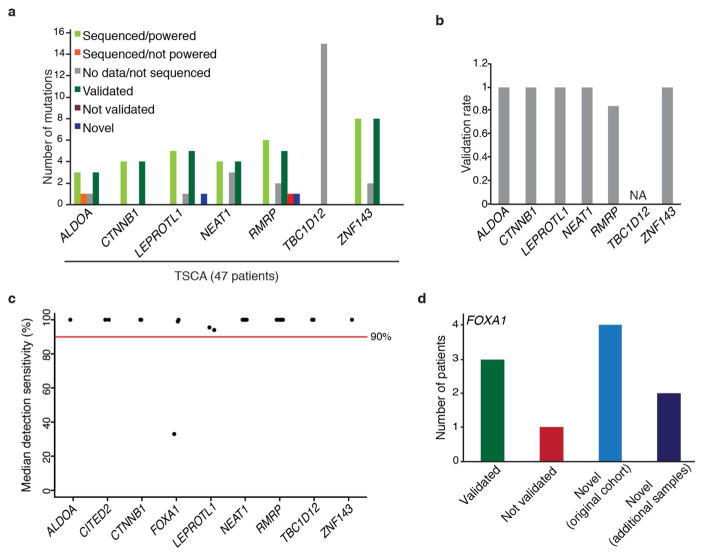
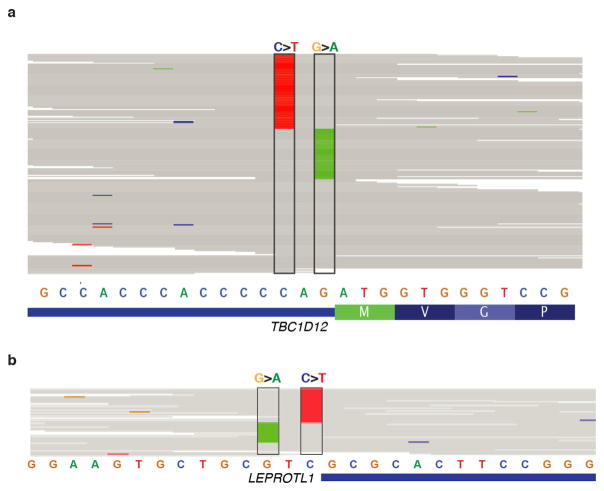
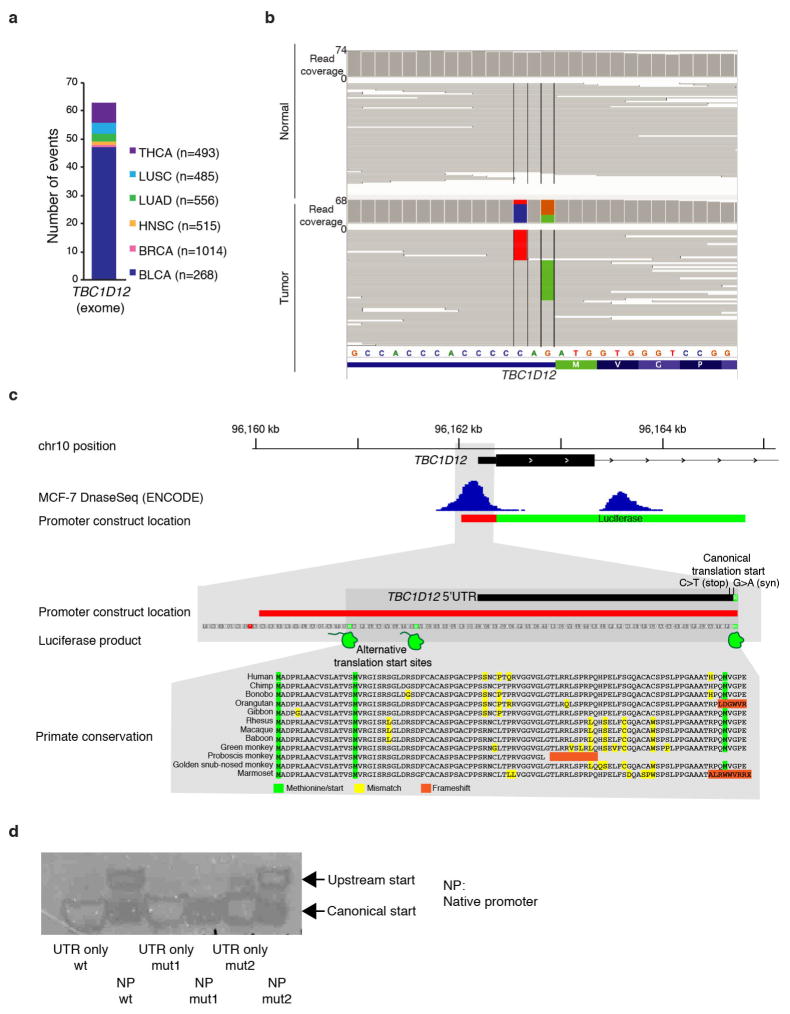
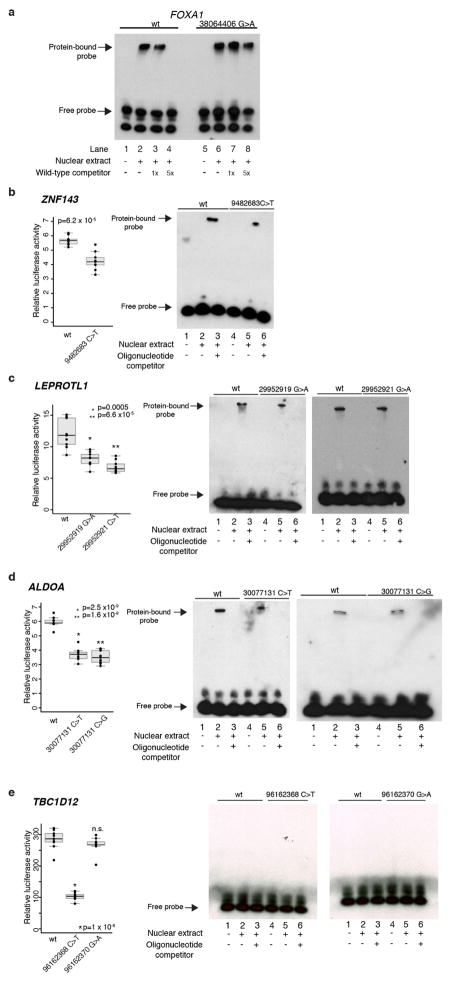
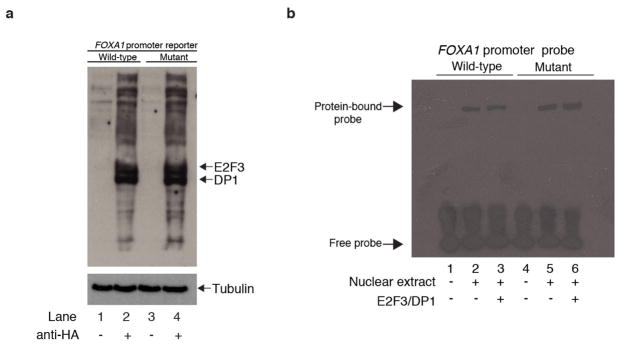
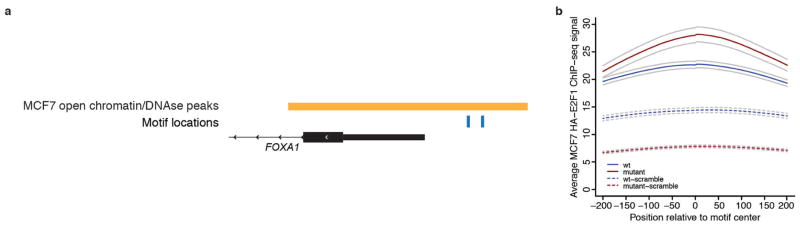
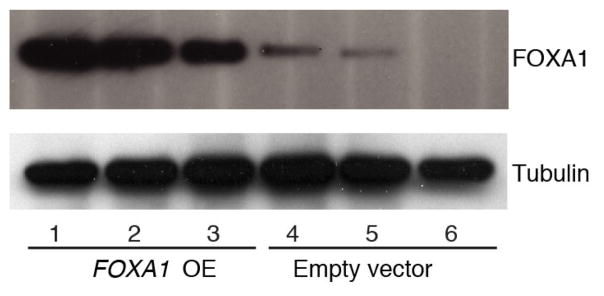
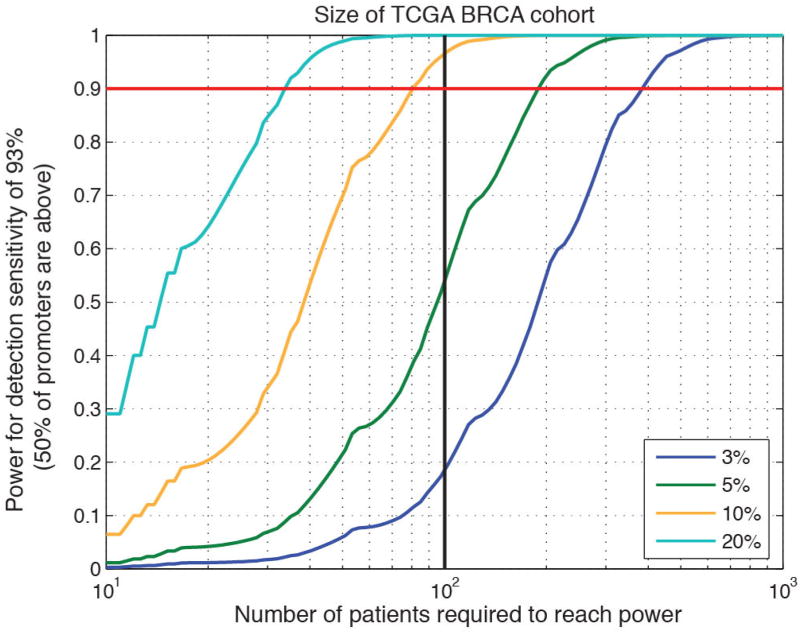
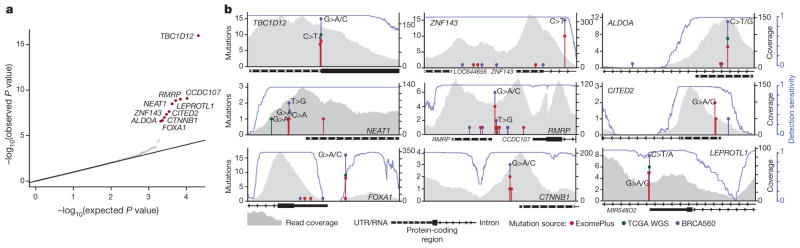
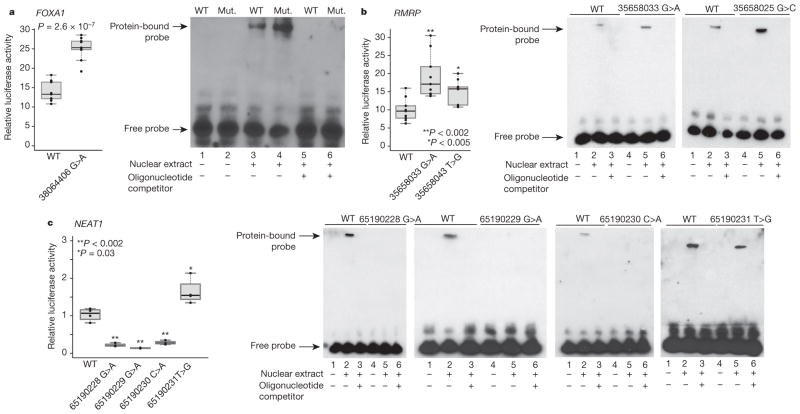
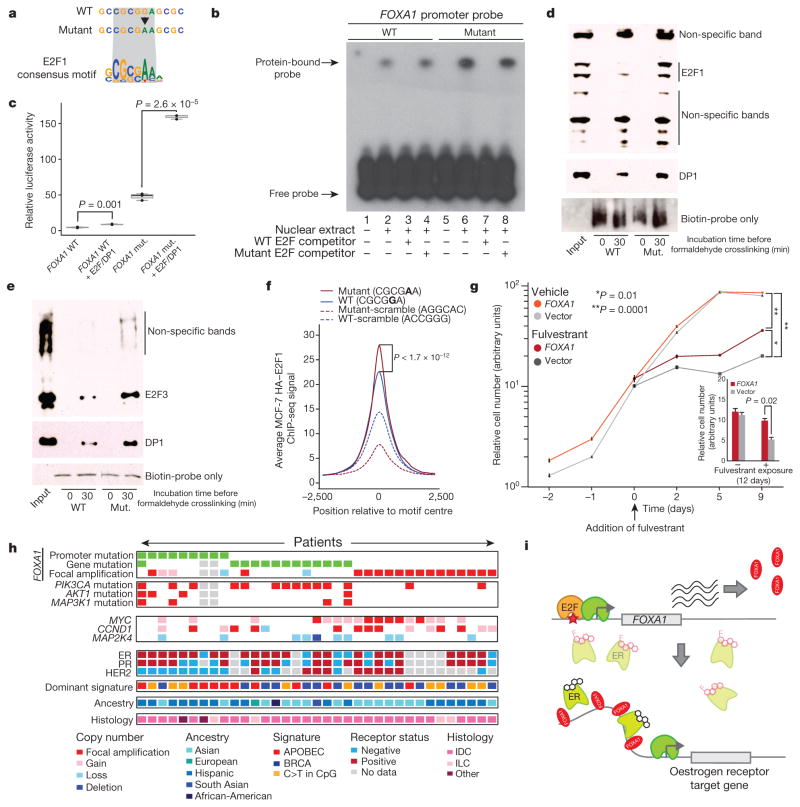
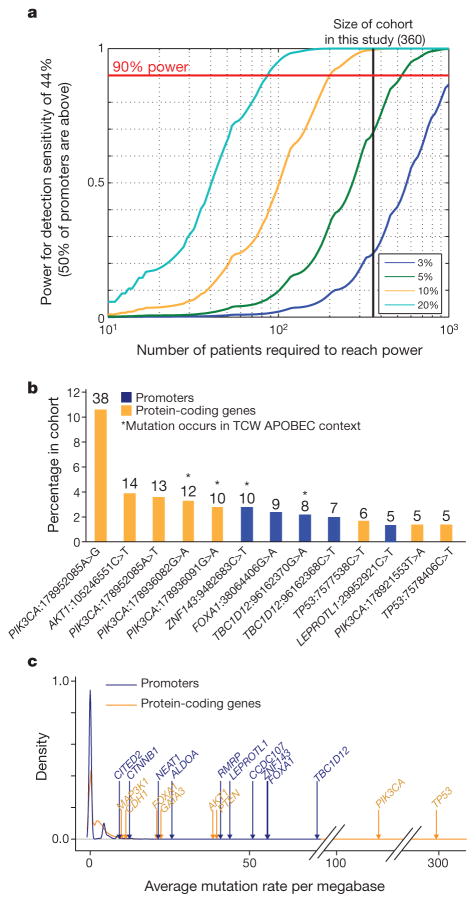
Genomic analysis of tumours has led to the identification of hundreds of cancer genes on the basis of the presence of mutations in protein-coding regions. By contrast, much less is known about cancer-causing mutations in non-coding regions. Here we perform deep sequencing in 360 primary breast cancers and develop computational methods to identify significantly mutated promoters. Clear signals are found in the promoters of three genes. FOXA1, a known driver of hormone-receptor positive breast cancer, harbours a mutational hotspot in its promoter leading to overexpression through increased E2F binding. RMRP and NEAT1, two non-coding RNA genes, carry mutations that affect protein binding to their promoters and alter expression levels. Our study shows that promoter regions harbour recurrent mutations in cancer with functional consequences and that the mutations occur at similar frequencies as in coding regions. Power analyses indicate that more such regions remain to be discovered through deep sequencing of adequately sized cohorts of patients.
Figures
Comment in
-
Cancer genomics: Less is more in the hunt for driver mutations.Nature. 2017 Jul 6;547(7661):40-41. doi: 10.1038/nature23085. Epub 2017 Jun 28. Nature. 2017. PMID: 28658210 No abstract available.
-
Absence of Promoter Mutation in TBC1D12 Gene in Solid and Hematologic Neoplasia.Pathol Oncol Res. 2019 Oct;25(4):1675-1676. doi: 10.1007/s12253-018-0468-7. Epub 2018 Sep 10. Pathol Oncol Res. 2019. PMID: 30203167 No abstract available.
-
Analysis of Promoter Mutation in Long Non-coding RNA NEAT1 in Acute Leukemias.Pathol Oncol Res. 2020 Apr;26(2):1345-1346. doi: 10.1007/s12253-019-00608-1. Epub 2019 Jan 29. Pathol Oncol Res. 2020. PMID: 30694517 No abstract available.
References
-
- Fredriksson NJ, Ny L, Nilsson JA, Larsson E. Systematic analysis of noncoding somatic mutations and gene expression alterations across 14 tumor types. Nat Genet. 2014;46:1258–1263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
