

One ligand, two regulators and three binding sites: How KDPG controls primary carbon metabolism in Pseudomonas
- PMID: 28658302
- PMCID: PMC5489143
- DOI: 10.1371/journal.pgen.1006839
One ligand, two regulators and three binding sites: How KDPG controls primary carbon metabolism in Pseudomonas
Abstract
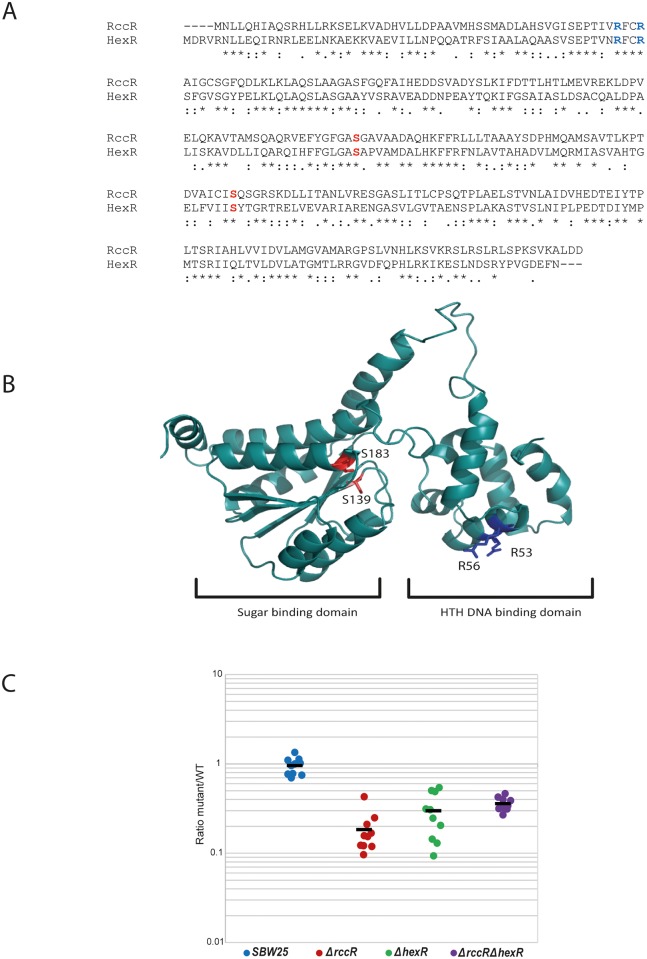
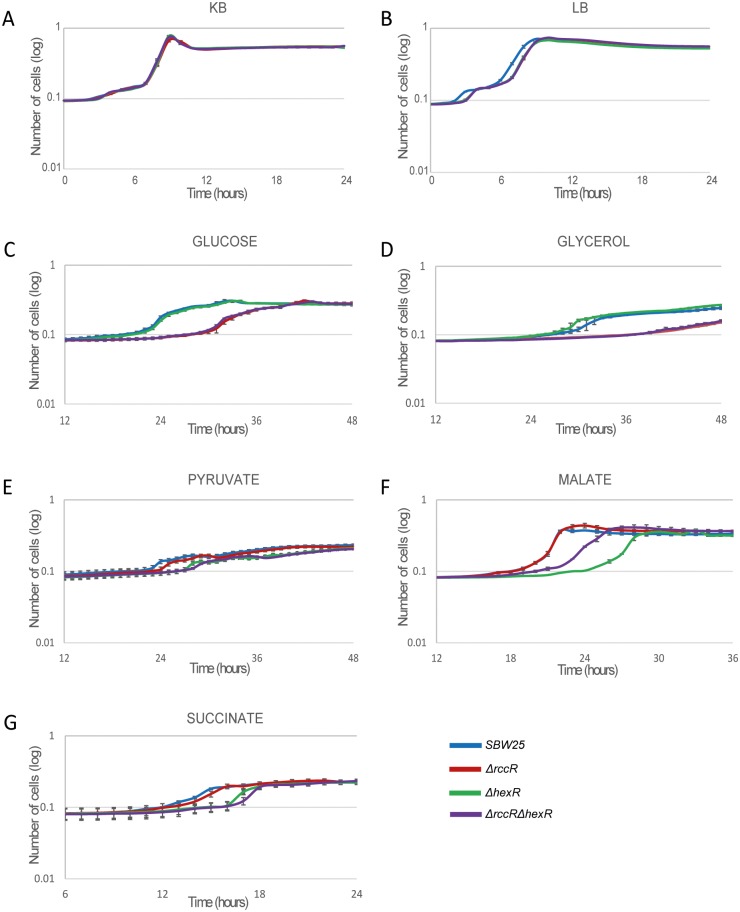
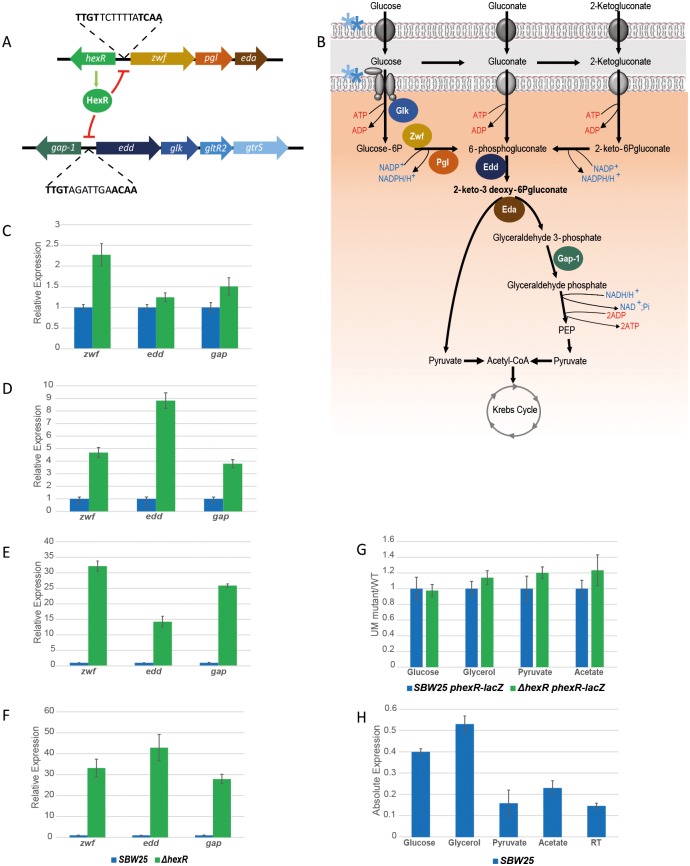
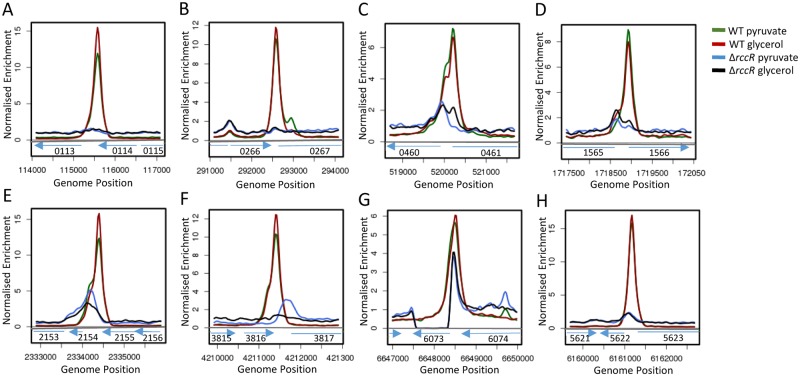
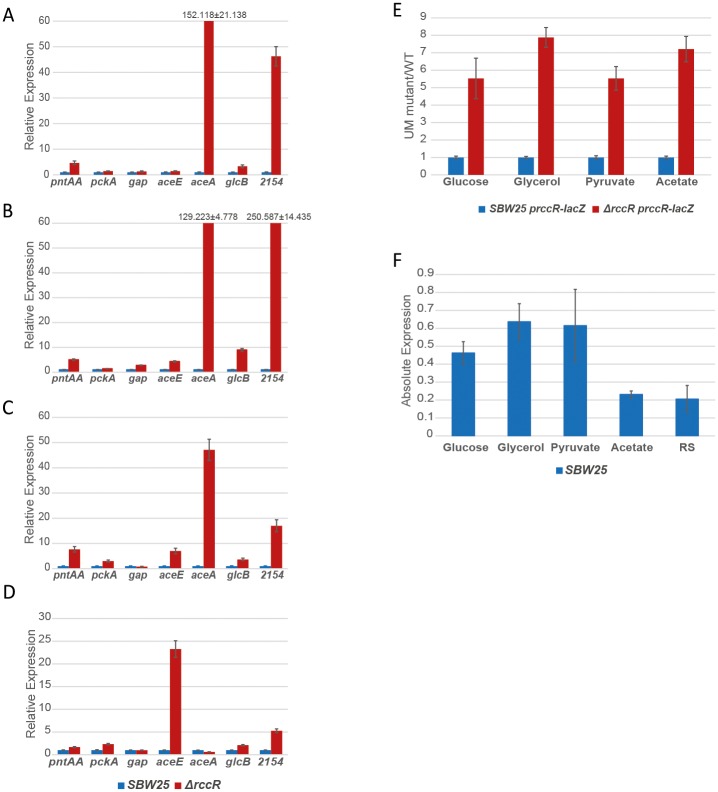
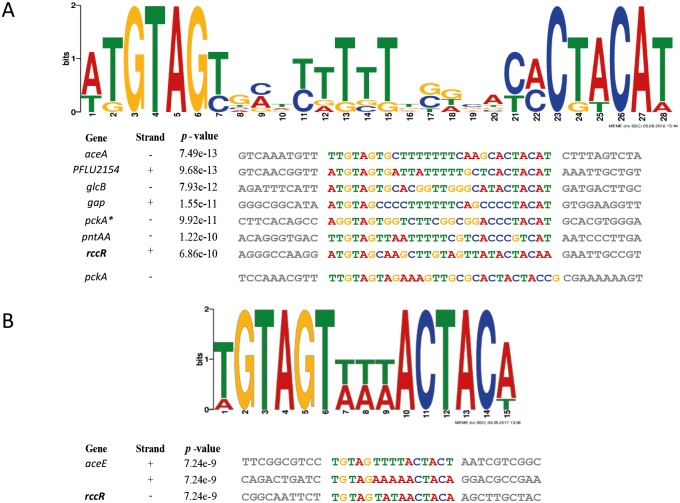
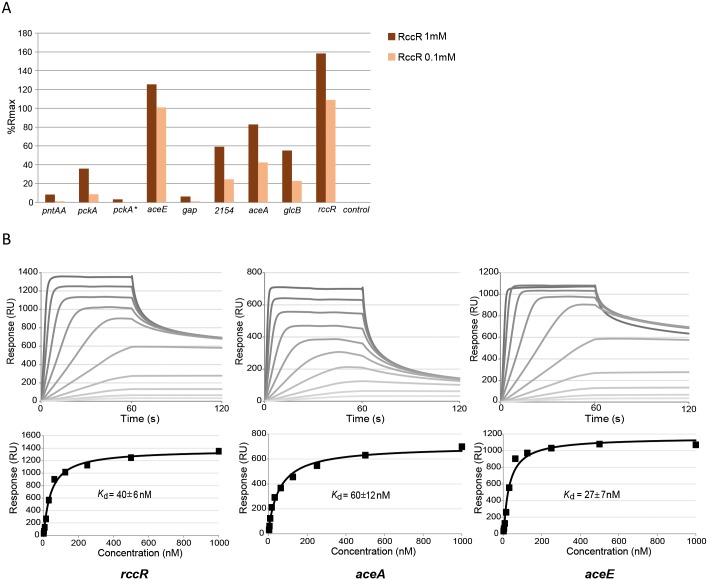
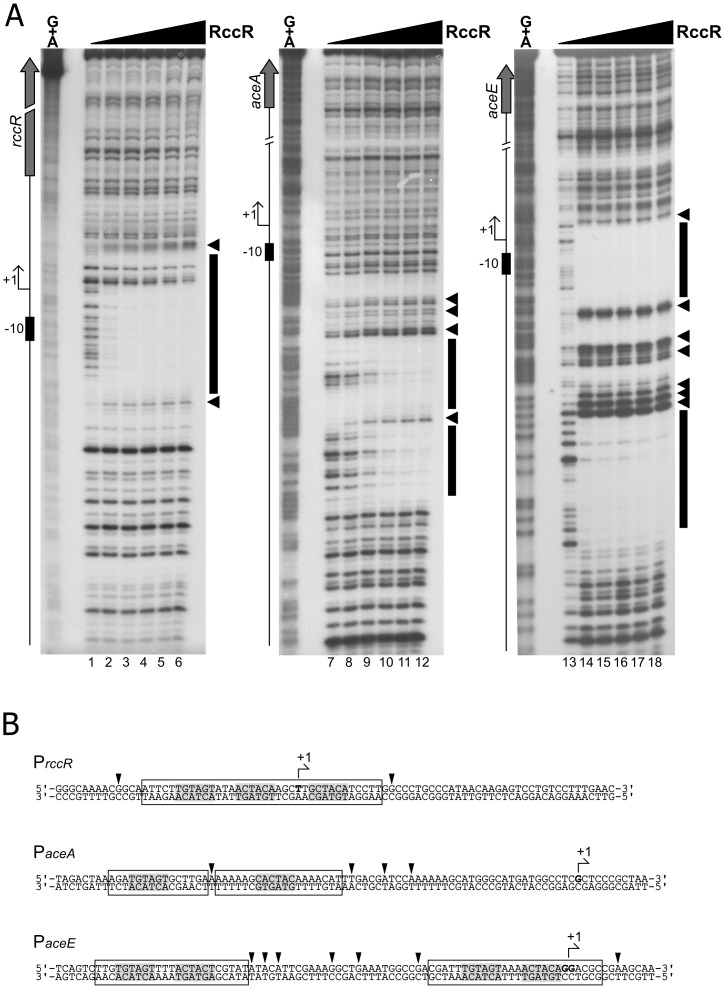
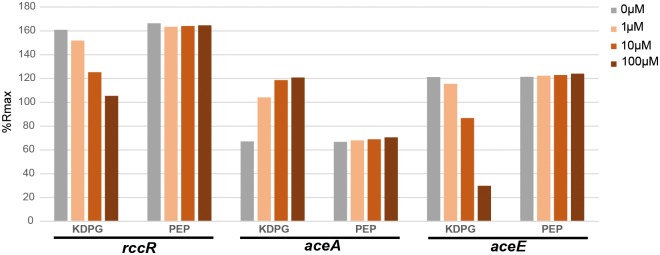
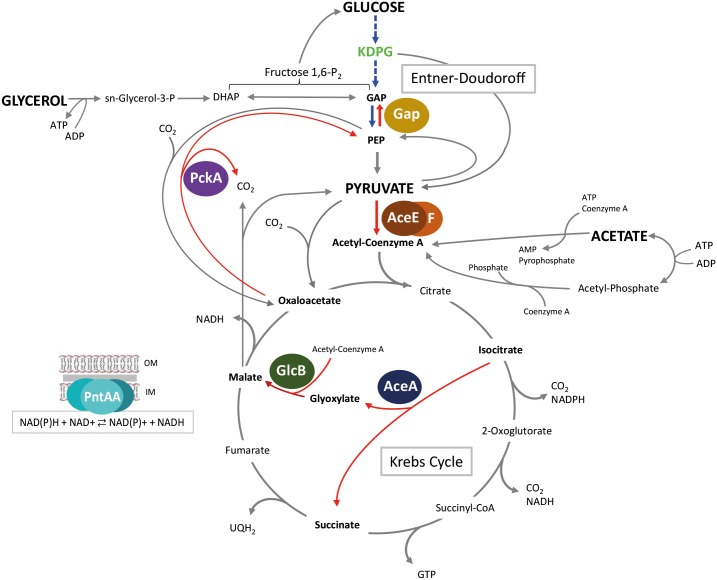
Effective regulation of primary carbon metabolism is critically important for bacteria to successfully adapt to different environments. We have identified an uncharacterised transcriptional regulator; RccR, that controls this process in response to carbon source availability. Disruption of rccR in the plant-associated microbe Pseudomonas fluorescens inhibits growth in defined media, and compromises its ability to colonise the wheat rhizosphere. Structurally, RccR is almost identical to the Entner-Doudoroff (ED) pathway regulator HexR, and both proteins are controlled by the same ED-intermediate; 2-keto-3-deoxy-6-phosphogluconate (KDPG). Despite these similarities, HexR and RccR control entirely different aspects of primary metabolism, with RccR regulating pyruvate metabolism (aceEF), the glyoxylate shunt (aceA, glcB, pntAA) and gluconeogenesis (pckA, gap). RccR displays complex and unusual regulatory behaviour; switching repression between the pyruvate metabolism and glyoxylate shunt/gluconeogenesis loci depending on the available carbon source. This regulatory complexity is enabled by two distinct pseudo-palindromic binding sites, differing only in the length of their linker regions, with KDPG binding increasing affinity for the 28 bp aceA binding site but decreasing affinity for the 15 bp aceE site. Thus, RccR is able to simultaneously suppress and activate gene expression in response to carbon source availability. Together, the RccR and HexR regulators enable the rapid coordination of multiple aspects of primary carbon metabolism, in response to levels of a single key intermediate.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Shimizu K. Metabolic Regulation and Coordination of the Metabolism in Bacteria in Response to a Variety of Growth Conditions. Adv Biochem Eng Biotechnol. 2016;155:1–54. doi: 10.1007/10_2015_320 . - DOI - PubMed
-
- Groisman EA, Mouslim C. Sensing by bacterial regulatory systems in host and non-host environments. Nat Rev Microbiol. 2006;4(9):705–9. doi: 10.1038/nrmicro1478 . - DOI - PubMed
-
- Silby MW, Winstanley C, Godfrey SA, Levy SB, Jackson RW. Pseudomonas genomes: diverse and adaptable. FEMS Microbiol Rev. 2011;35(4):652–80. . - PubMed
-
- Little RH, Grenga L, Saalbach G, Howat AM, Pfeilmeier S, Trampari E, et al. Adaptive Remodeling of the Bacterial Proteome by Specific Ribosomal Modification Regulates Pseudomonas Infection and Niche Colonisation. PLoS Genet. 2016;12(2):e1005837 doi: 10.1371/journal.pgen.1005837 . - DOI - PMC - PubMed
-
- Barahona E, Navazo A, Yousef-Coronado F, Aguirre de Carcer D, Martinez-Granero F, Espinosa-Urgel M, et al. Efficient rhizosphere colonization by Pseudomonas fluorescens f113 mutants unable to form biofilms on abiotic surfaces. Environ Microbiol. 2010;12(12):3185–95. Epub 2010/07/16. doi: 10.1111/j.1462-2920.2010.02291.x . - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
