

Definition of a High-Confidence Mitochondrial Proteome at Quantitative Scale
- PMID: 28658629
- PMCID: PMC5494306
- DOI: 10.1016/j.celrep.2017.06.014
Definition of a High-Confidence Mitochondrial Proteome at Quantitative Scale
Abstract
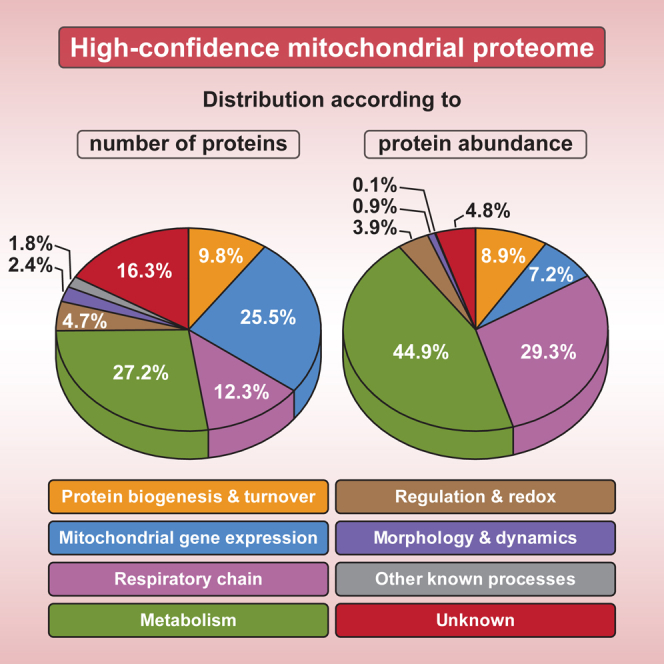
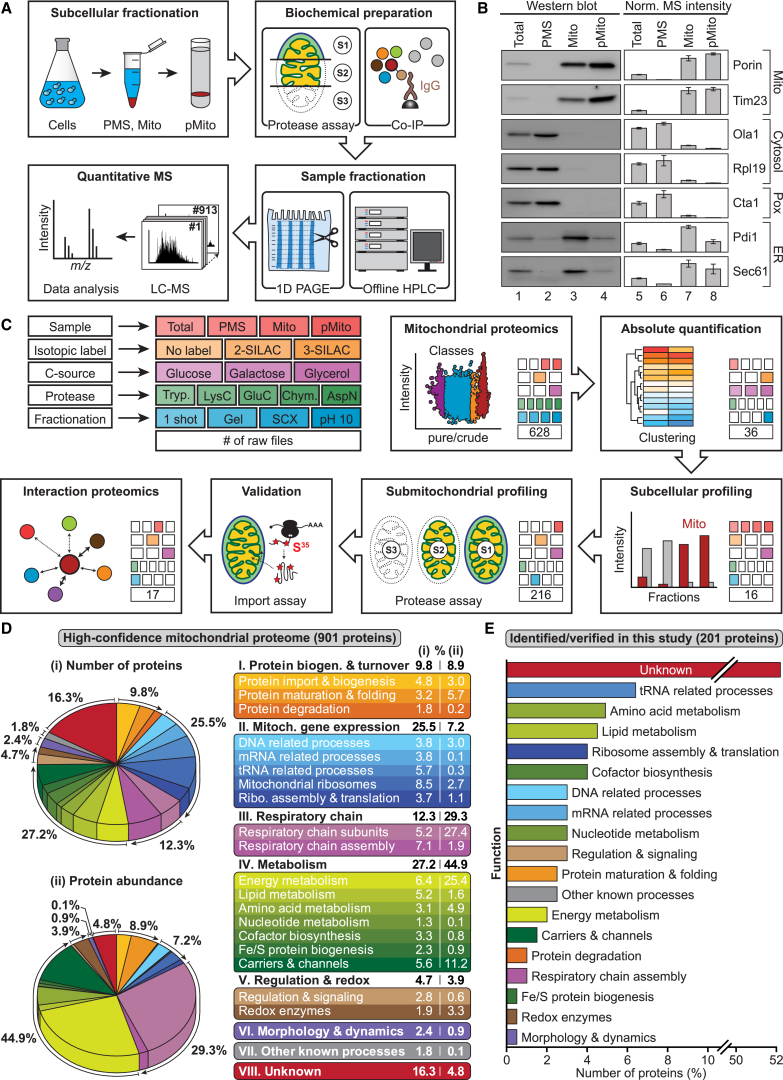
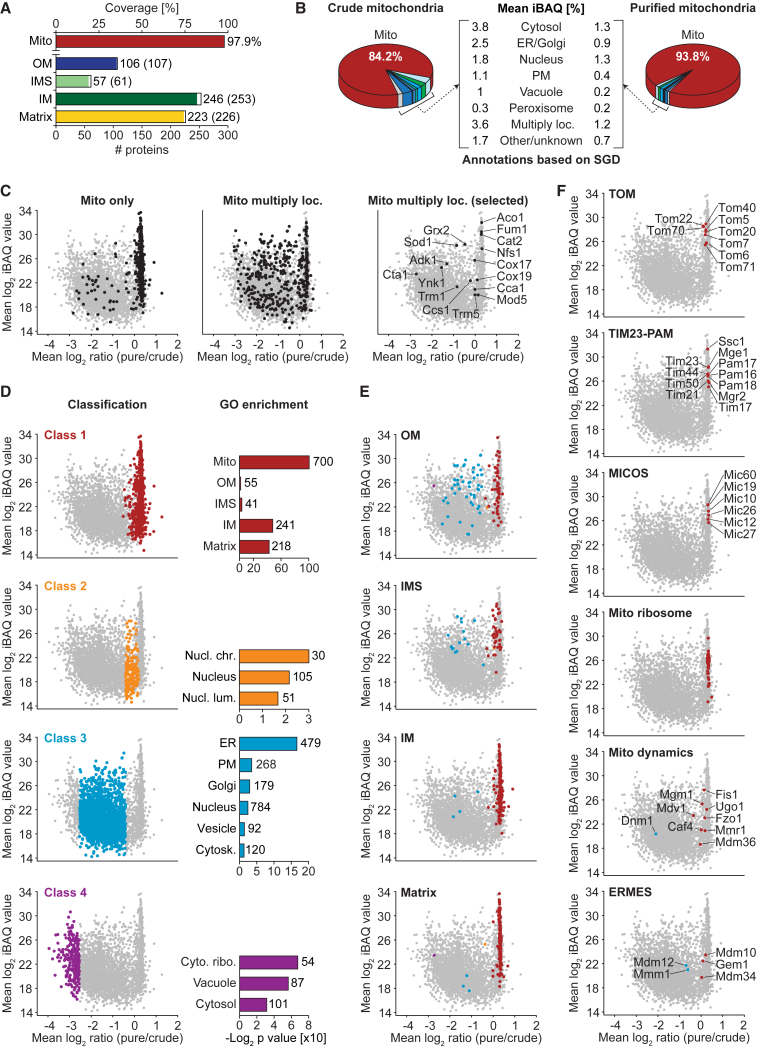
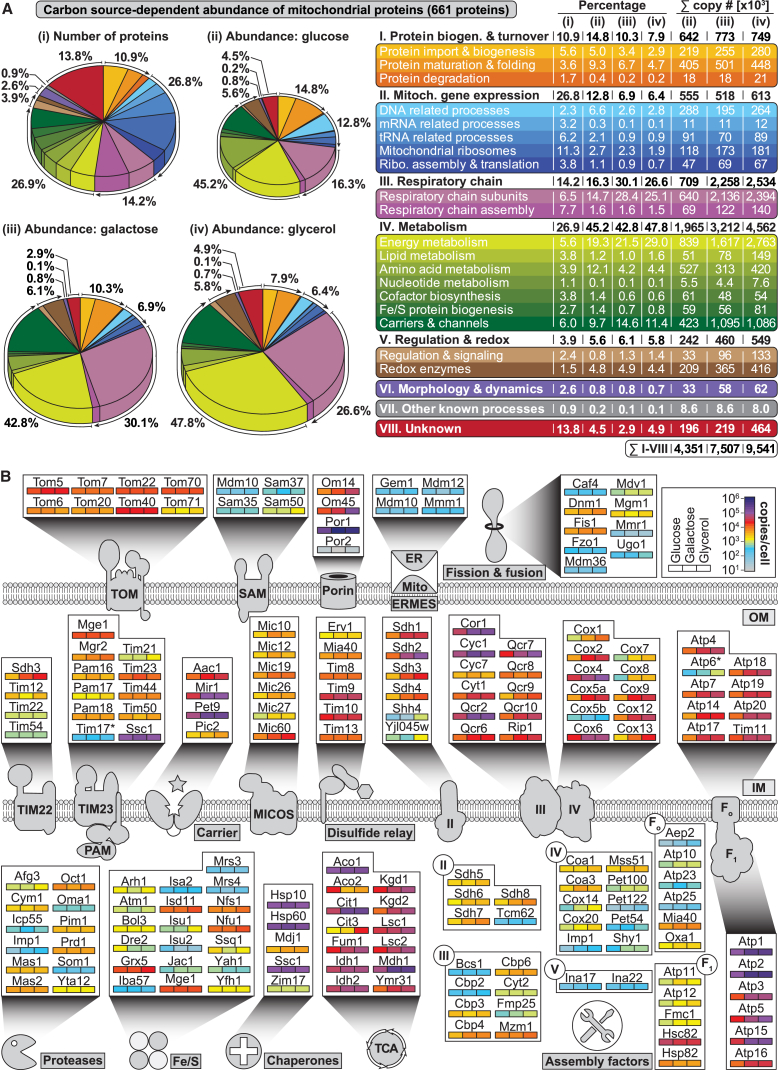
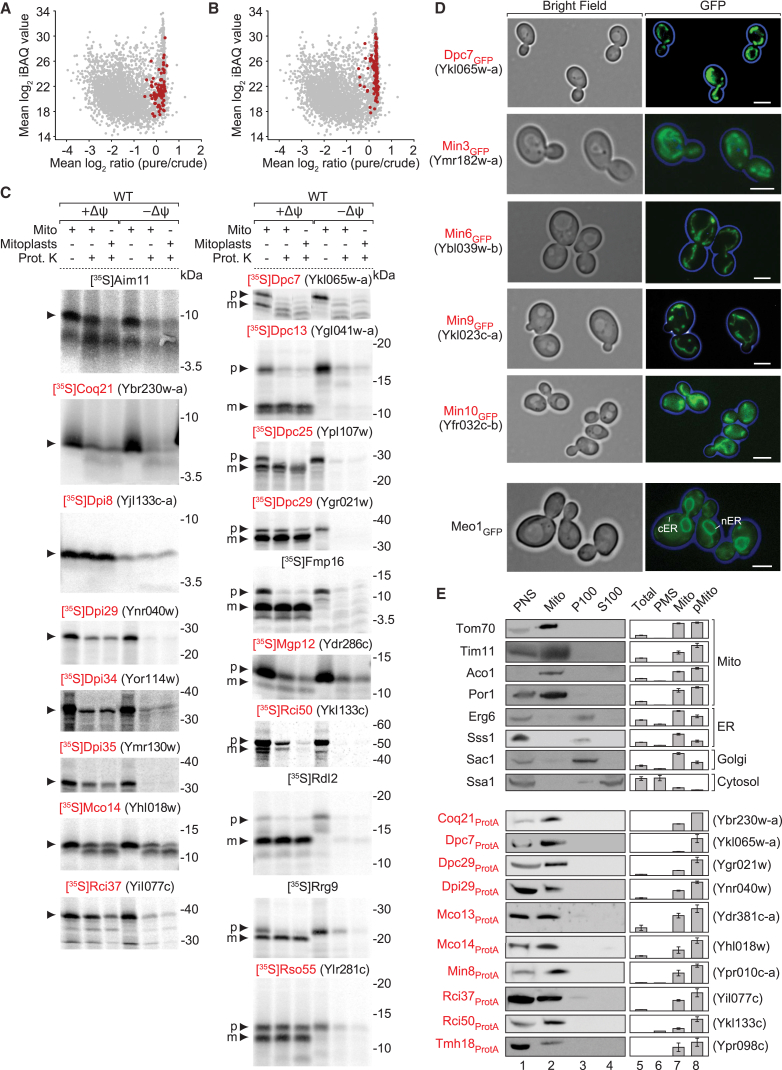
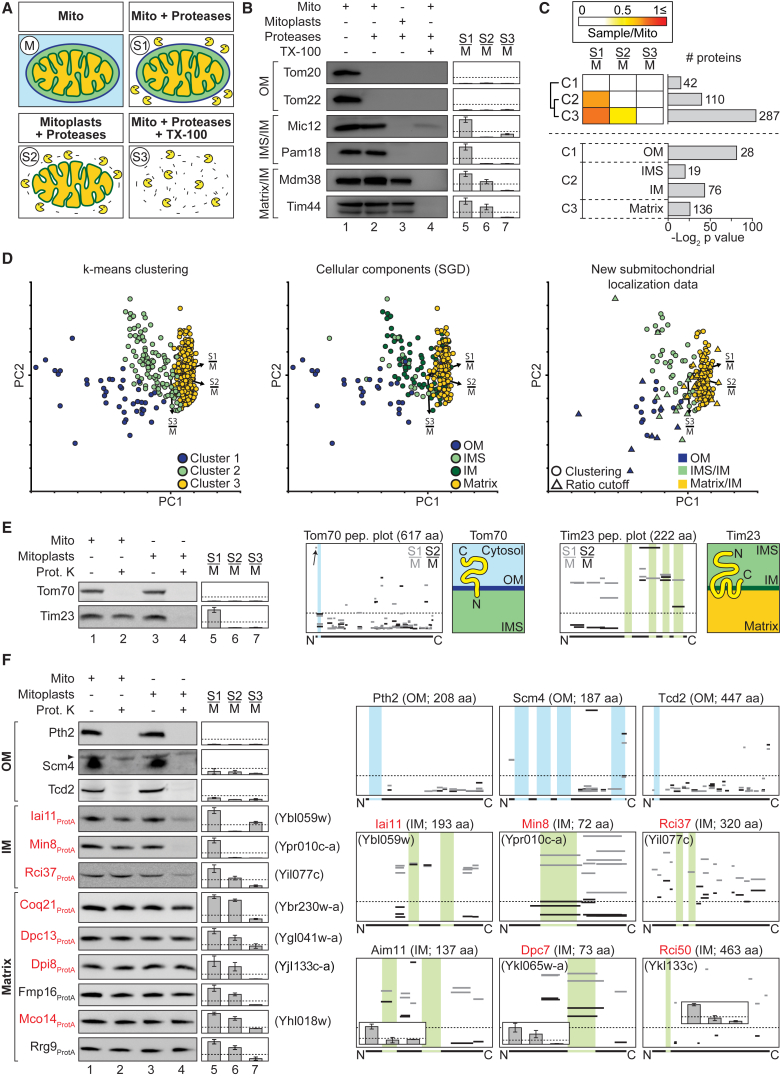
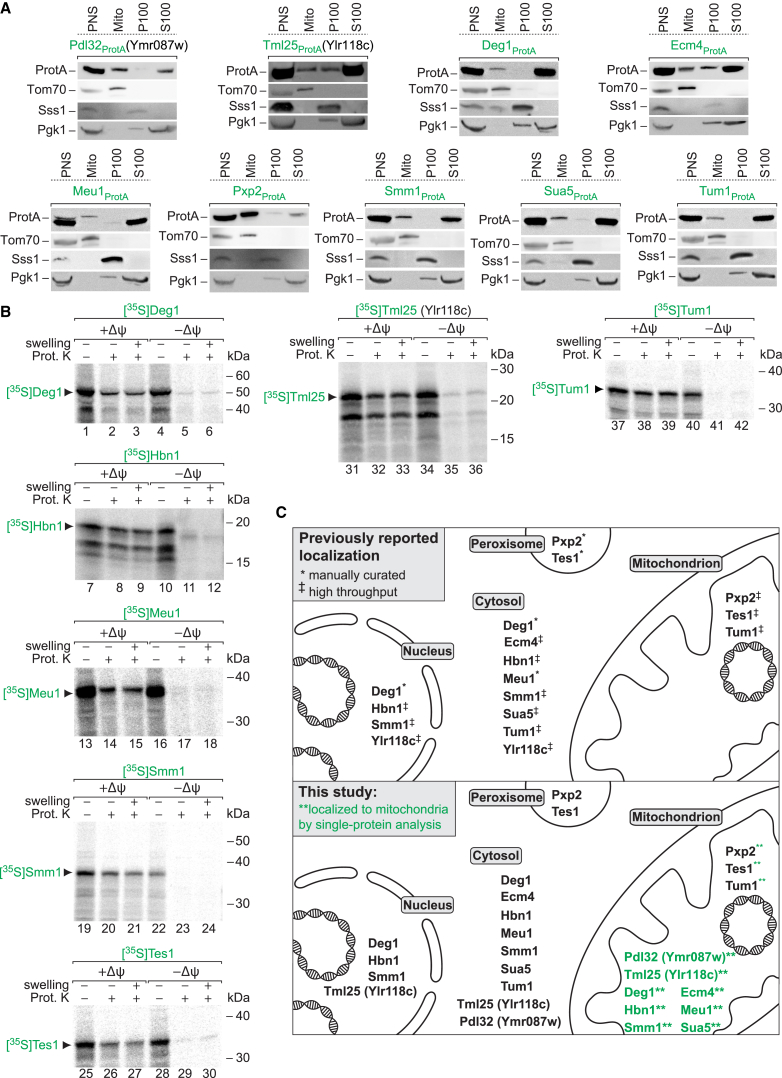
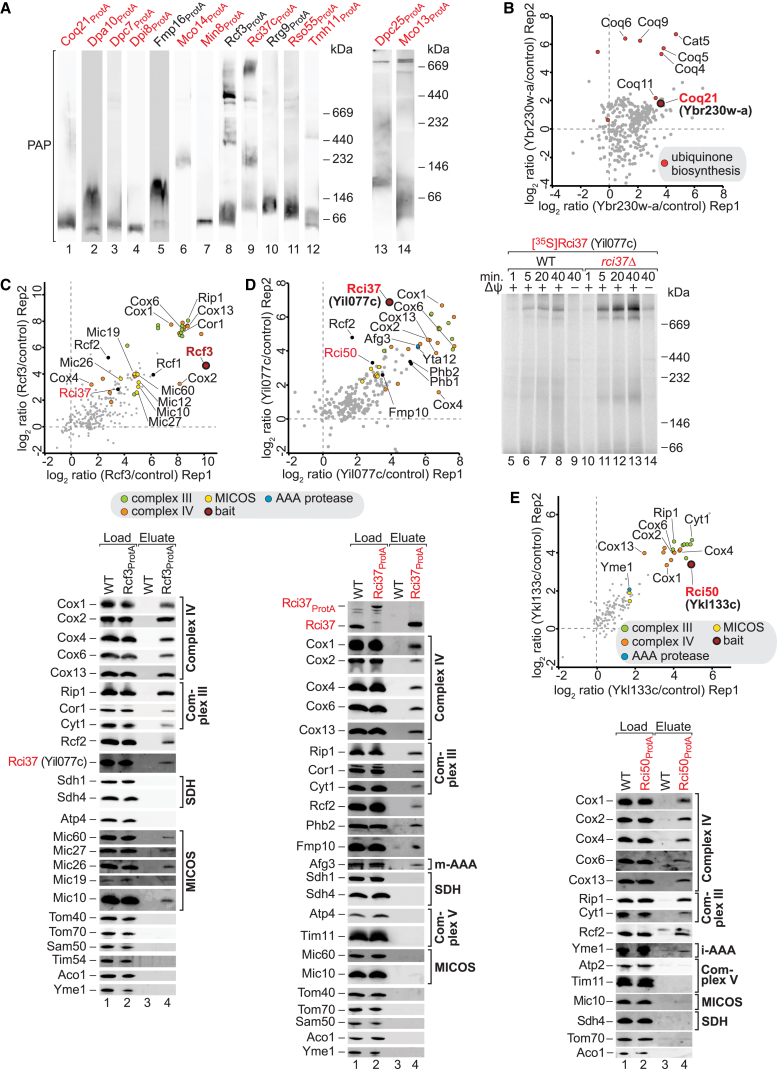
Mitochondria perform central functions in cellular bioenergetics, metabolism, and signaling, and their dysfunction has been linked to numerous diseases. The available studies cover only part of the mitochondrial proteome, and a separation of core mitochondrial proteins from associated fractions has not been achieved. We developed an integrative experimental approach to define the proteome of yeast mitochondria. We classified > 3,300 proteins of mitochondria and mitochondria-associated fractions and defined 901 high-confidence mitochondrial proteins, expanding the set of mitochondrial proteins by 82. Our analysis includes protein abundance under fermentable and nonfermentable growth, submitochondrial localization, single-protein experiments, and subcellular classification of mitochondria-associated fractions. We identified mitochondrial interactors of respiratory chain supercomplexes, ATP synthase, AAA proteases, the mitochondrial contact site and cristae organizing system (MICOS), and the coenzyme Q biosynthesis cluster, as well as mitochondrial proteins with dual cellular localization. The integrative proteome provides a high-confidence source for the characterization of physiological and pathophysiological functions of mitochondria and their integration into the cellular environment.
Keywords: SILAC; dual localization; mitochondria; protein copy numbers; protein-protein interaction; quantitative MS; subcellular localization; submitochondrial localization.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Alder N.N., Jensen R.E., Johnson A.E. Fluorescence mapping of mitochondrial TIM23 complex reveals a water-facing, substrate-interacting helix surface. Cell. 2008;134:439–450. - PubMed
-
- Barbot M., Jans D.C., Schulz C., Denkert N., Kroppen B., Hoppert M., Jakobs S., Meinecke M. Mic10 oligomerizes to bend mitochondrial inner membranes at cristae junctions. Cell Metab. 2015;21:756–763. - PubMed
-
- Bohnert M., Zerbes R.M., Davies K.M., Mühleip A.W., Rampelt H., Horvath S.E., Boenke T., Kram A., Perschil I., Veenhuis M. Central role of Mic10 in the mitochondrial contact site and cristae organizing system. Cell Metab. 2015;21:747–755. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
