

Differential Expression of VEGF-Axxx Isoforms Is Critical for Development of Pulmonary Fibrosis
- PMID: 28661183
- PMCID: PMC5564672
- DOI: 10.1164/rccm.201603-0568OC
Differential Expression of VEGF-Axxx Isoforms Is Critical for Development of Pulmonary Fibrosis
Abstract
Rationale: Fibrosis after lung injury is related to poor outcome, and idiopathic pulmonary fibrosis (IPF) can be regarded as an exemplar. Vascular endothelial growth factor (VEGF)-A has been implicated in this context, but there are conflicting reports as to whether it is a contributory or protective factor. Differential splicing of the VEGF-A gene produces multiple functional isoforms including VEGF-A165a and VEGF-A165b, a member of the inhibitory family. To date there is no clear information on the role of VEGF-A in IPF.
Objectives: To establish VEGF-A isoform expression and functional effects in IPF.
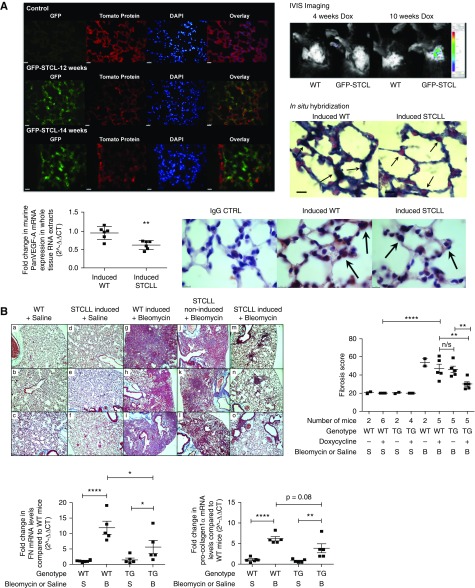
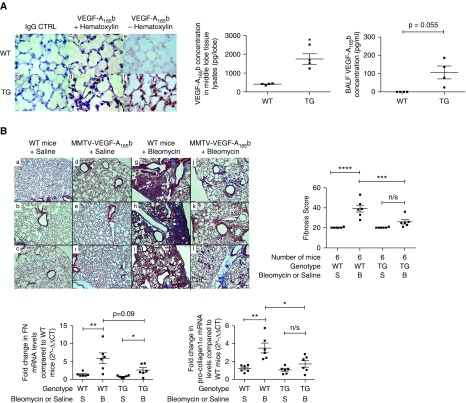
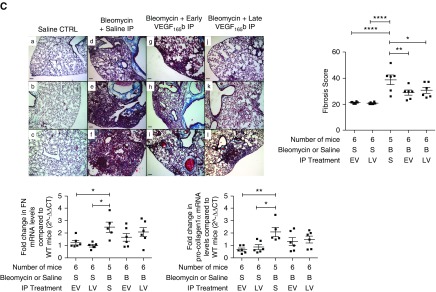
Methods: We used tissue sections, plasma, and lung fibroblasts from patients with IPF and control subjects. In a bleomycin-induced lung fibrosis model we used wild-type MMTV mice and a triple transgenic mouse SPC-rtTA+/-TetoCre+/-LoxP-VEGF-A+/+ to conditionally induce VEGF-A isoform deletion specifically in the alveolar type II (ATII) cells of adult mice.
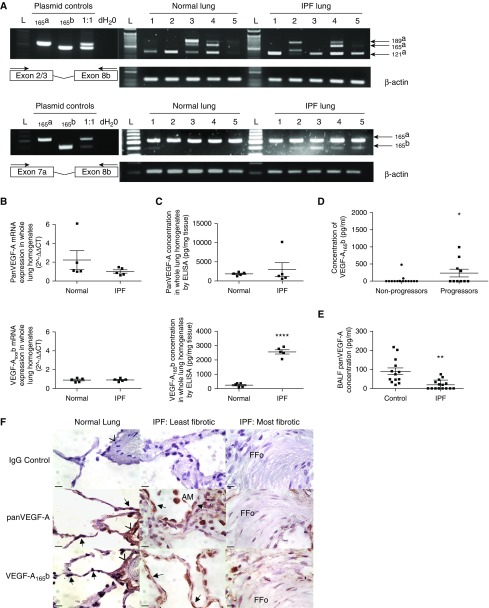
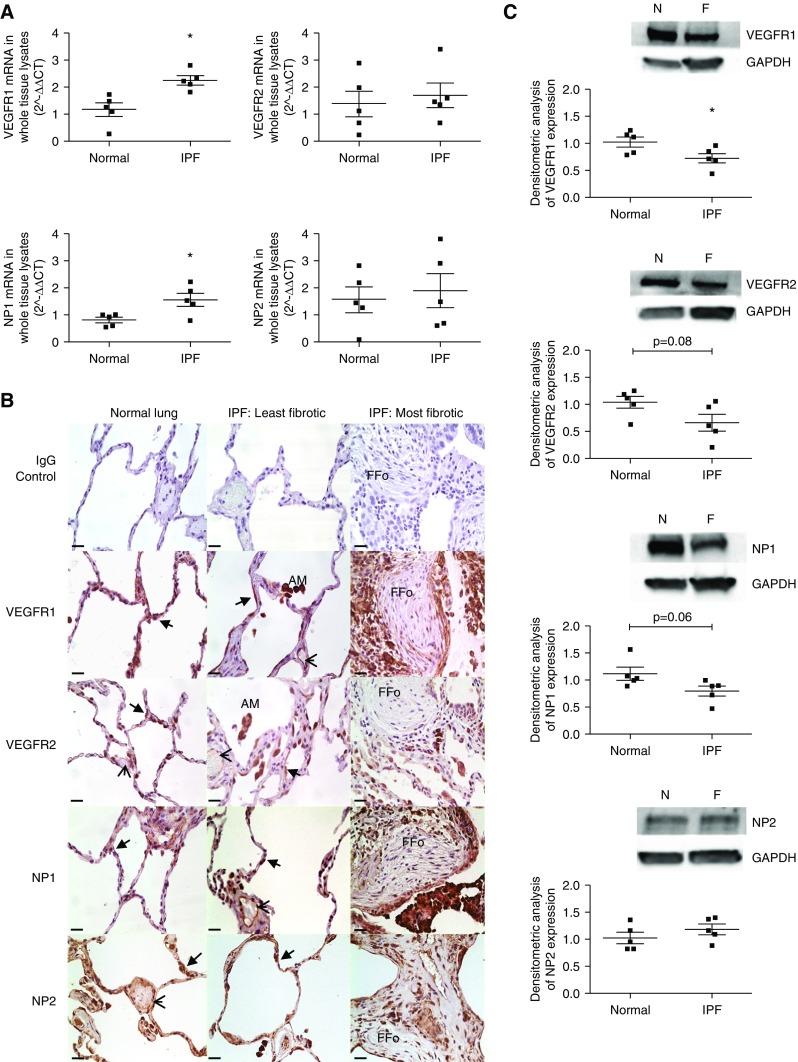
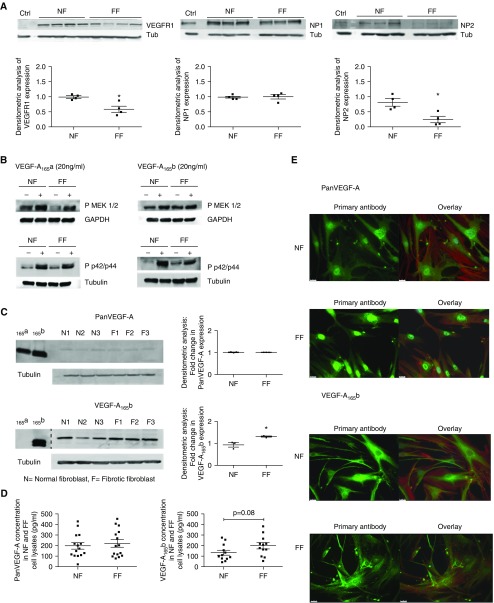
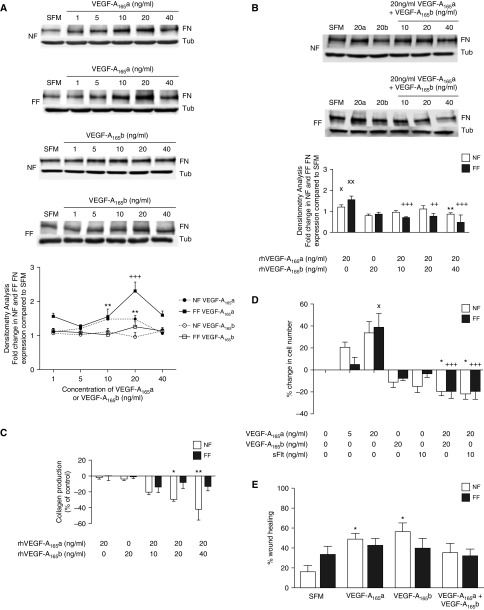
Measurements and main results: IPF and normal lung fibroblasts differentially expressed and responded to VEGF-A165a and VEGF-A165b in terms of proliferation and matrix expression. Increased VEGF-A165b was detected in plasma of progressing patients with IPF. In a mouse model of pulmonary fibrosis, ATII-specific deficiency of VEGF-A or constitutive overexpression of VEGF-A165b inhibited the development of pulmonary fibrosis, as did treatment with intraperitoneal delivery of VEGF-A165b to wild-type mice.
Conclusions: These results indicate that changes in the bioavailability of VEGF-A sourced from ATII cells, namely the ratio of VEGF-Axxxa to VEGF-Axxxb, are critical in development of pulmonary fibrosis and may be a paradigm for the regulation of tissue repair.
Keywords: animal models of pulmonary fibrosis; idiopathic pulmonary fibrosis; vascular endothelial growth factor.
Figures
Comment in
-
Vascular Endothelial Growth Factor in Idiopathic Pulmonary Fibrosis. An Imbalancing Act.Am J Respir Crit Care Med. 2017 Aug 15;196(4):409-411. doi: 10.1164/rccm.201703-0605ED. Am J Respir Crit Care Med. 2017. PMID: 28809513 No abstract available.
References
-
- Navaratnam V, Fleming KM, West J, Smith CJ, Jenkins RG, Fogarty A, Hubbard RB. The rising incidence of idiopathic pulmonary fibrosis in the U.K. Thorax. 2011;66:462–467. - PubMed
-
- Collard HR, King TE, Jr, Bartelson BB, Vourlekis JS, Schwarz MI, Brown KK. Changes in clinical and physiologic variables predict survival in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 2003;168:538–542. - PubMed
-
- Raghu G, Weycker D, Edelsberg J, Bradford WZ, Oster G. Incidence and prevalence of idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 2006;174:810–816. - PubMed
-
- Olson AL, Swigris JJ, Lezotte DC, Norris JM, Wilson CG, Brown KK. Mortality from pulmonary fibrosis increased in the United States from 1992 to 2003. Am J Respir Crit Care Med. 2007;176:277–284. - PubMed
-
- Ferrara N. Vascular endothelial growth factor: basic science and clinical progress. Endocr Rev. 2004;25:581–611. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
