

Structural basis for human respiratory syncytial virus NS1-mediated modulation of host responses
- PMID: 28665409
- PMCID: PMC5739881
- DOI: 10.1038/nmicrobiol.2017.101
Structural basis for human respiratory syncytial virus NS1-mediated modulation of host responses
Abstract
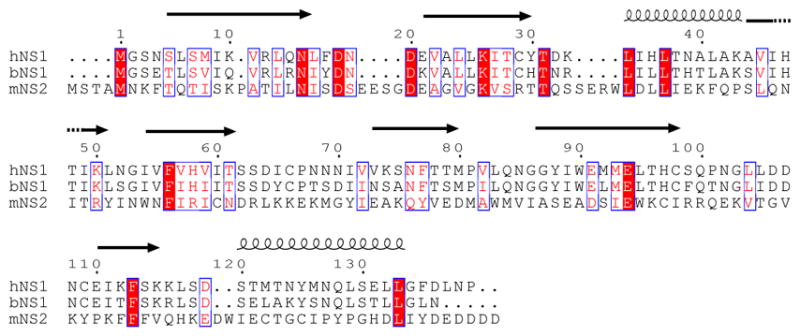
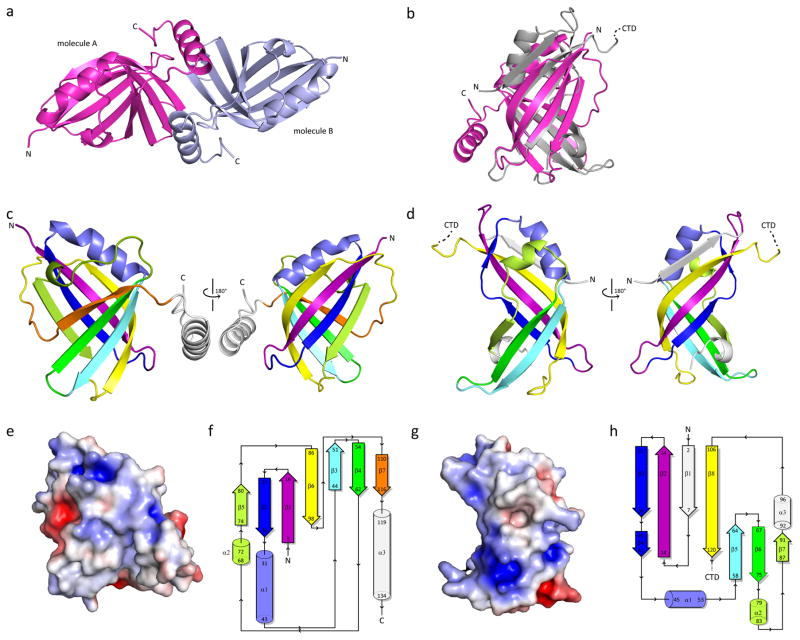
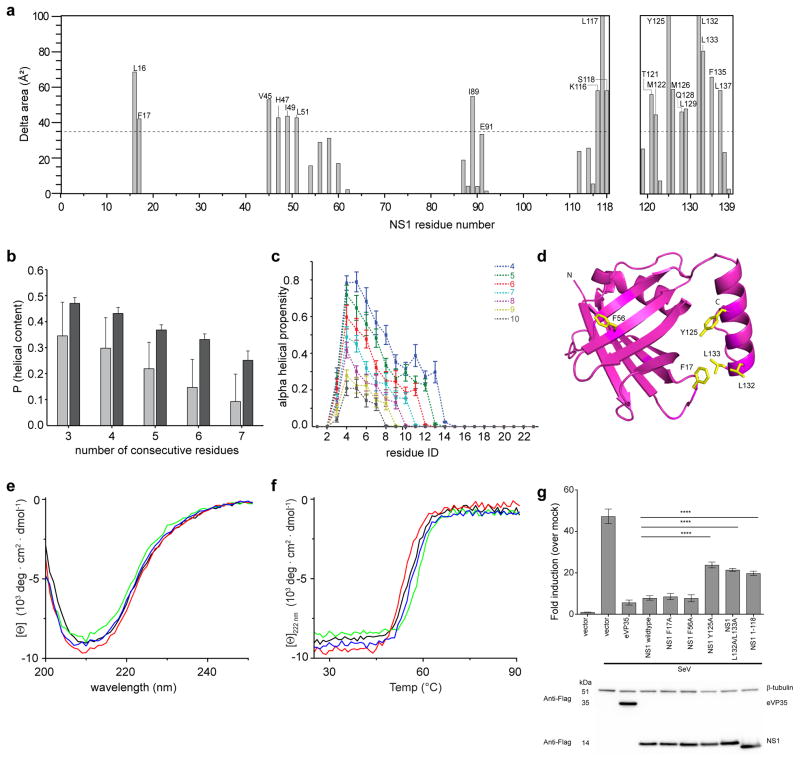
Human respiratory syncytial virus (hRSV) is a major cause of morbidity and mortality in the paediatric, elderly and immune-compromised populations1,2. A gap in our understanding of hRSV disease pathology is the interplay between virally encoded immune antagonists and host components that limit hRSV replication. hRSV encodes for non-structural (NS) proteins that are important immune antagonists3-6; however, the role of these proteins in viral pathogenesis is incompletely understood. Here, we report the crystal structure of hRSV NS1 protein, which suggests that NS1 is a structural paralogue of hRSV matrix (M) protein. Comparative analysis of the shared structural fold with M revealed regions unique to NS1. Studies on NS1 wild type or mutant alone or in recombinant RSVs demonstrate that structural regions unique to NS1 contribute to modulation of host responses, including inhibition of type I interferon responses, suppression of dendritic cell maturation and promotion of inflammatory responses. Transcriptional profiles of A549 cells infected with recombinant RSVs show significant differences in multiple host pathways, suggesting that NS1 may have a greater role in regulating host responses than previously appreciated. These results provide a framework to target NS1 for therapeutic development to limit hRSV-associated morbidity and mortality.
Figures
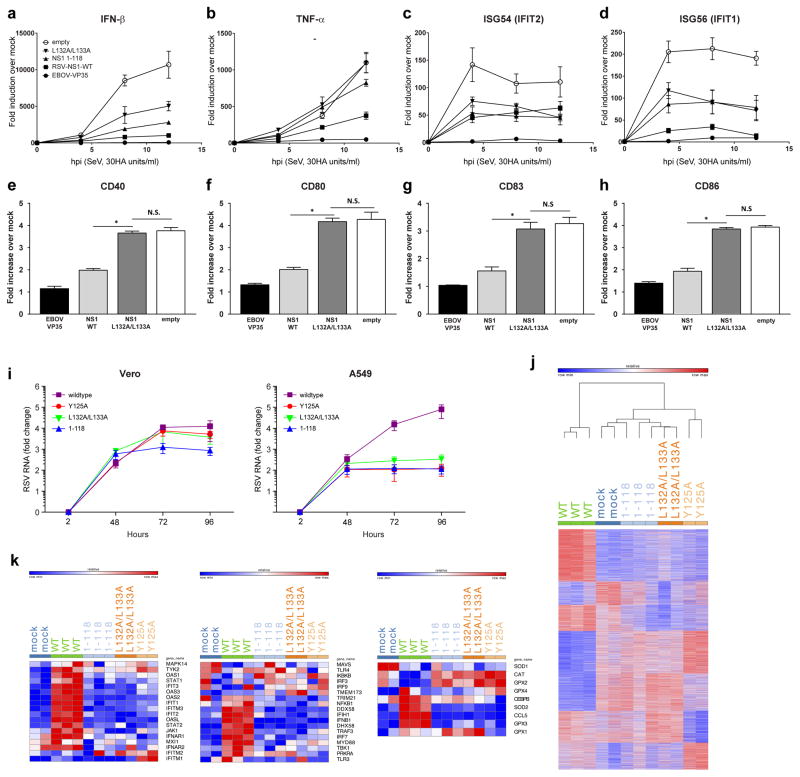
 , NS1 wildtype;
, NS1 wildtype;
 , NS1 1-118;
, NS1 1-118;
 , NS1 Y125A; and
, NS1 Y125A; and
 , NS1 L132A/L133A. j, Heatmap representing genes with number of matching reads greater than five in A549 cells 96 hpi post infection with hRSV NS1 wildtype or mutant proteins. Colors represent expression values; red indicates higher expression and blue indicates lower expression within a gene expression profile. k, Heatmaps representing the members of the top set of genes in IFN induction (left panel), IFN response (middle panel), and antioxidative response (right panel) signaling pathways in A549 cells 96 hpi with hRSV NS1 wildtype or mutant proteins.
, NS1 L132A/L133A. j, Heatmap representing genes with number of matching reads greater than five in A549 cells 96 hpi post infection with hRSV NS1 wildtype or mutant proteins. Colors represent expression values; red indicates higher expression and blue indicates lower expression within a gene expression profile. k, Heatmaps representing the members of the top set of genes in IFN induction (left panel), IFN response (middle panel), and antioxidative response (right panel) signaling pathways in A549 cells 96 hpi with hRSV NS1 wildtype or mutant proteins.References
-
- Hall CB. The burgeoning burden of respiratory syncytial virus among children. Infectious disorders drug targets. 2012;12:92–97. - PubMed
-
- Stockman LJ, Curns AT, Anderson LJ, Fischer-Langley G. Respiratory syncytial virus-associated hospitalizations among infants and young children in the United States, 1997–2006. The Pediatric infectious disease journal. 2012;31:5–9. - PubMed
MeSH terms
Substances
Grants and funding
- R01 AI087798/AI/NIAID NIH HHS/United States
- R01 AI125453/AI/NIAID NIH HHS/United States
- R01 AI107056/AI/NIAID NIH HHS/United States
- U19 AI070489/AI/NIAID NIH HHS/United States
- R01 AI111605/AI/NIAID NIH HHS/United States
- T32 CA009547/CA/NCI NIH HHS/United States
- U19 AI109945/AI/NIAID NIH HHS/United States
- R01 AI123926/AI/NIAID NIH HHS/United States
- P01 AI120943/AI/NIAID NIH HHS/United States
- R01 AI114654/AI/NIAID NIH HHS/United States
- U19 AI095227/AI/NIAID NIH HHS/United States
- R01 AI130591/AI/NIAID NIH HHS/United States
- UH3 HL123429/HL/NHLBI NIH HHS/United States
- U19 AI109664/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
