

The PKR regulatory subunit of protein kinase A (PKA) is involved in the regulation of growth, sexual and asexual development, and pathogenesis in Fusarium graminearum
- PMID: 28665481
- PMCID: PMC6638095
- DOI: 10.1111/mpp.12576
The PKR regulatory subunit of protein kinase A (PKA) is involved in the regulation of growth, sexual and asexual development, and pathogenesis in Fusarium graminearum
Abstract
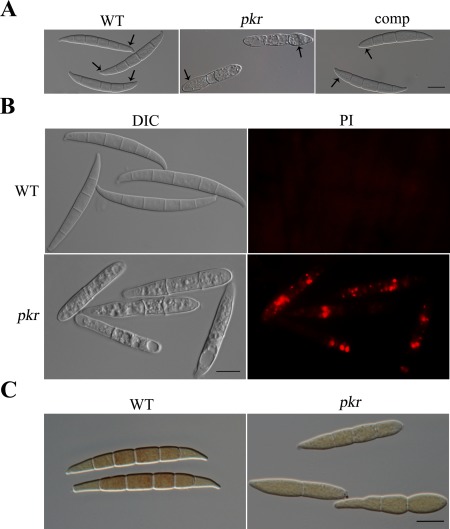
Fusarium graminearum is a causal agent of wheat scab disease and a producer of deoxynivalenol (DON) mycotoxins. Treatment with exogenous cyclic adenosine monophosphate (cAMP) increases its DON production. In this study, to better understand the role of the cAMP-protein kinase A (PKA) pathway in F. graminearum, we functionally characterized the PKR gene encoding the regulatory subunit of PKA. Mutants deleted of PKR were viable, but showed severe defects in growth, conidiation and plant infection. The pkr mutant produced compact colonies with shorter aerial hyphae with an increased number of nuclei in hyphal compartments. Mutant conidia were morphologically abnormal and appeared to undergo rapid autophagy-related cell death. The pkr mutant showed blocked perithecium development, but increased DON production. It had a disease index of less than unity and failed to spread to neighbouring spikelets. The mutant was unstable and spontaneous suppressors with a faster growth rate were often produced on older cultures. A total of 67 suppressor strains that grew faster than the original mutant were isolated. Three showed a similar growth rate and colony morphology to the wild-type, but were still defective in conidiation. Sequencing analysis with 18 candidate PKA-related genes in three representative suppressor strains identified mutations only in the CPK1 catalytic subunit gene. Further characterization showed that 10 of the other 64 suppressor strains also had mutations in CPK1. Overall, these results showed that PKR is important for the regulation of hyphal growth, reproduction, pathogenesis and DON production, and mutations in CPK1 are partially suppressive to the deletion of PKR in F. graminearum.
Keywords: DON; autophagy; conidiation; fungal pathogenicity; suppressor.
© 2017 BSPP AND JOHN WILEY & SONS LTD.
Figures






References
-
- Alexander, N.J. , Proctor, R.H. and McCormick, S.P. (2009) Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium . Toxin Rev. 28, 198–215.
-
- Bai, G.H. and Shaner, G. (2004) Management and resistance in wheat and barley to Fusarium head blight. Annu. Rev. Phytopathol. 42, 135–161. - PubMed
-
- Bai, G.H. , Desjardins, A.E. and Plattner, R.D. (2002) Deoxynivalenol‐nonproducing Fusarium graminearum causes initial infection, but does not cause disease spread in wheat spikes. Mycopathologia, 153, 91–98. - PubMed
-
- Bluhm, B.H. , Zhao, X. , Flaherty, J.E. , Xu, J.R. and Dunkle, L.D. (2007) RAS2 regulates growth and pathogenesis in Fusarium graminearum . Mol. Plant–Microbe Interact. 20, 627–636. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
