

Type 2 Diabetes Variants Disrupt Function of SLC16A11 through Two Distinct Mechanisms
- PMID: 28666119
- PMCID: PMC5562285
- DOI: 10.1016/j.cell.2017.06.011
Type 2 Diabetes Variants Disrupt Function of SLC16A11 through Two Distinct Mechanisms
Abstract
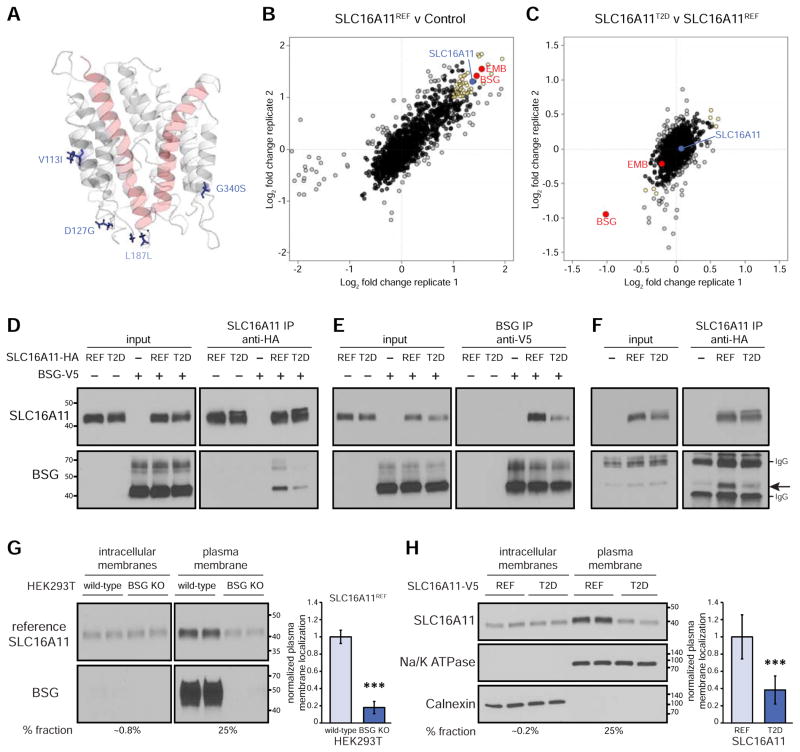
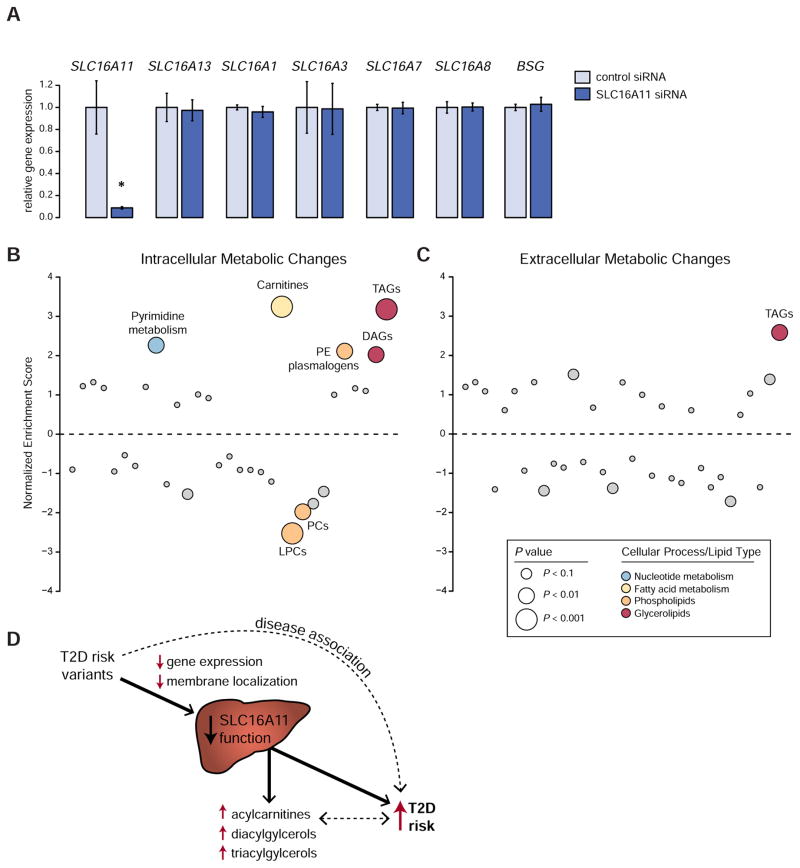
Type 2 diabetes (T2D) affects Latinos at twice the rate seen in populations of European descent. We recently identified a risk haplotype spanning SLC16A11 that explains ∼20% of the increased T2D prevalence in Mexico. Here, through genetic fine-mapping, we define a set of tightly linked variants likely to contain the causal allele(s). We show that variants on the T2D-associated haplotype have two distinct effects: (1) decreasing SLC16A11 expression in liver and (2) disrupting a key interaction with basigin, thereby reducing cell-surface localization. Both independent mechanisms reduce SLC16A11 function and suggest SLC16A11 is the causal gene at this locus. To gain insight into how SLC16A11 disruption impacts T2D risk, we demonstrate that SLC16A11 is a proton-coupled monocarboxylate transporter and that genetic perturbation of SLC16A11 induces changes in fatty acid and lipid metabolism that are associated with increased T2D risk. Our findings suggest that increasing SLC16A11 function could be therapeutically beneficial for T2D. VIDEO ABSTRACT.
Keywords: MCT11; SLC16A11; disease mechanism; fatty acid metabolism; genetics; lipid metabolism; monocarboxylates; precision medicine; solute carrier (SLC); type 2 diabetes (T2D).
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
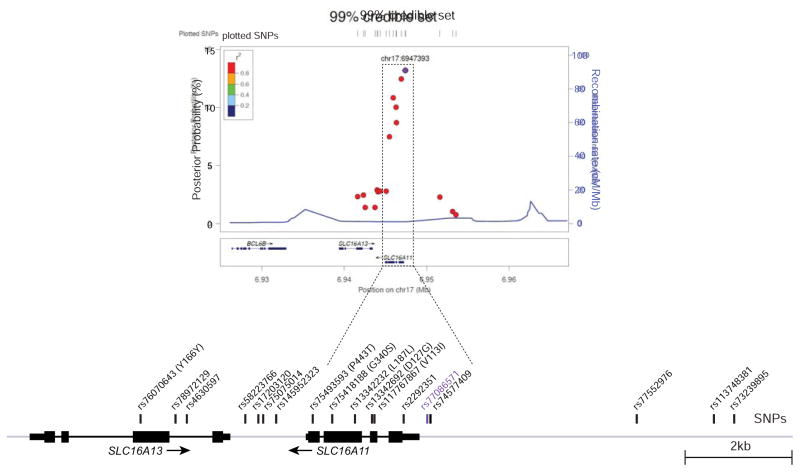
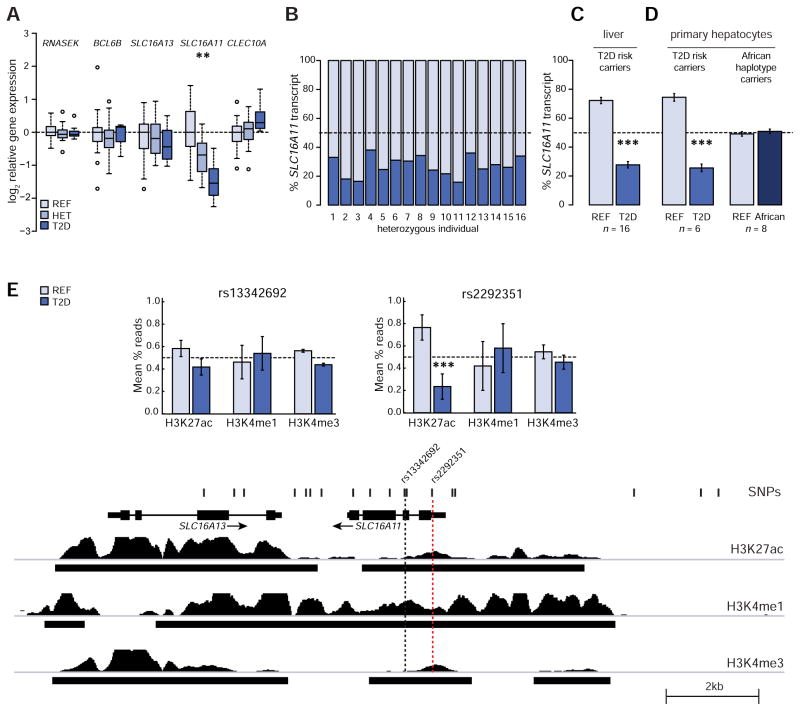
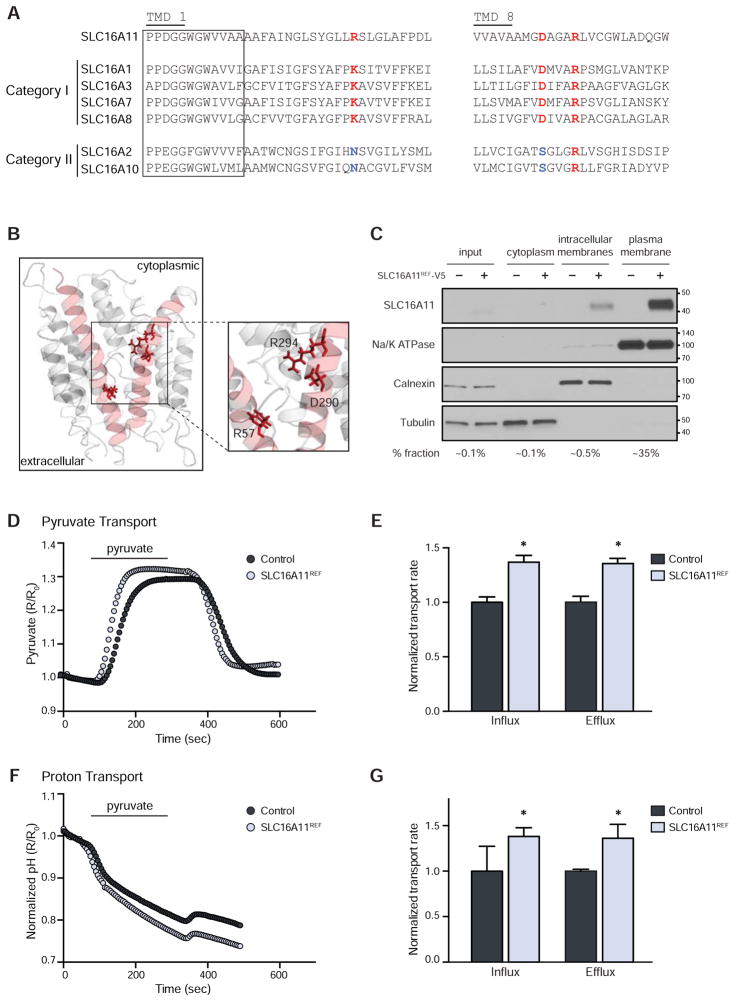
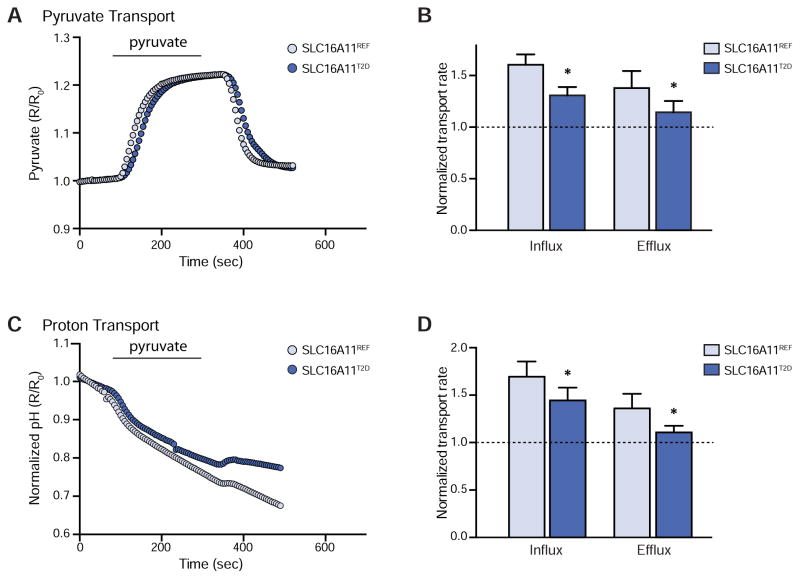
Comment in
-
A New Drug Target for Type 2 Diabetes.Cell. 2017 Jun 29;170(1):12-14. doi: 10.1016/j.cell.2017.06.024. Cell. 2017. PMID: 28666114
References
-
- Adams SH, Hoppel CL, Lok KH, Zhao L, Wong SW, Minkler PE, Hwang DH, Newman JW, Garvey WT. Plasma acylcarnitine profiles suggest incomplete long-chain fatty acid beta-oxidation and altered tricarboxylic acid cycle activity in type 2 diabetic African-American women. J Nutr. 2009;139:1073–1081. - PMC - PubMed
-
- Bernstein BE, Kamal M, Lindblad-Toh K, Bekiranov S, Bailey DK, Huebert DJ, McMahon S, Karlsson EK, Kulbokas EJ, 3rd, Gingeras TR, et al. Genomic maps and comparative analysis of histone modifications in human and mouse. Cell. 2005;120:169–181. - PubMed
-
- Cohen JC, Boerwinkle E, Mosley TH, Jr, Hobbs HH. Sequence variations in PCSK9, low LDL, and protection against coronary heart disease. N Engl J Med. 2006;354:1264–1272. - PubMed
-
- Delaneau O, Zagury JF, Marchini J. Improved whole-chromosome phasing for disease and population genetic studies. Nat Methods. 2013;10:5–6. - PubMed
-
- Diamond J. The double puzzle of diabetes. Nature. 2003;423:599–602. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
