

The methyltransferase SETDB1 regulates a large neuron-specific topological chromatin domain
- PMID: 28671686
- PMCID: PMC5560095
- DOI: 10.1038/ng.3906
The methyltransferase SETDB1 regulates a large neuron-specific topological chromatin domain
Abstract
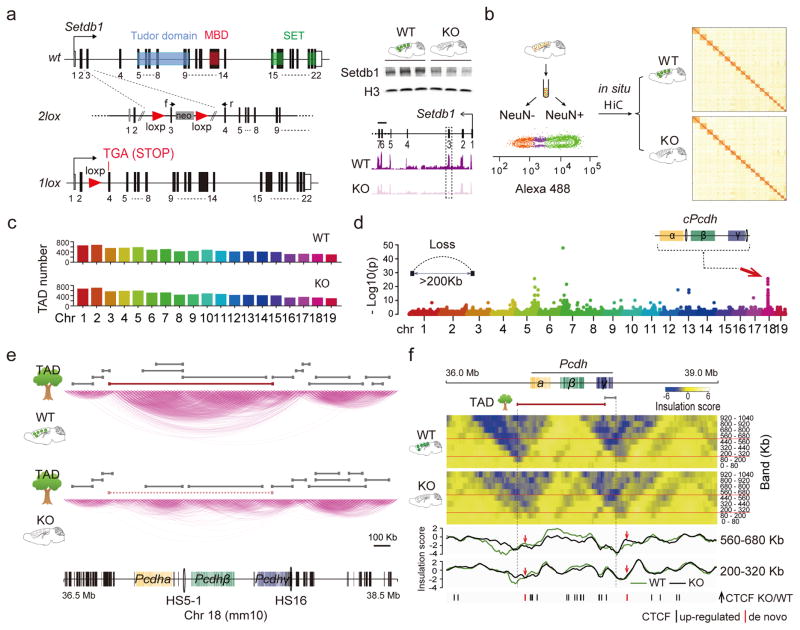
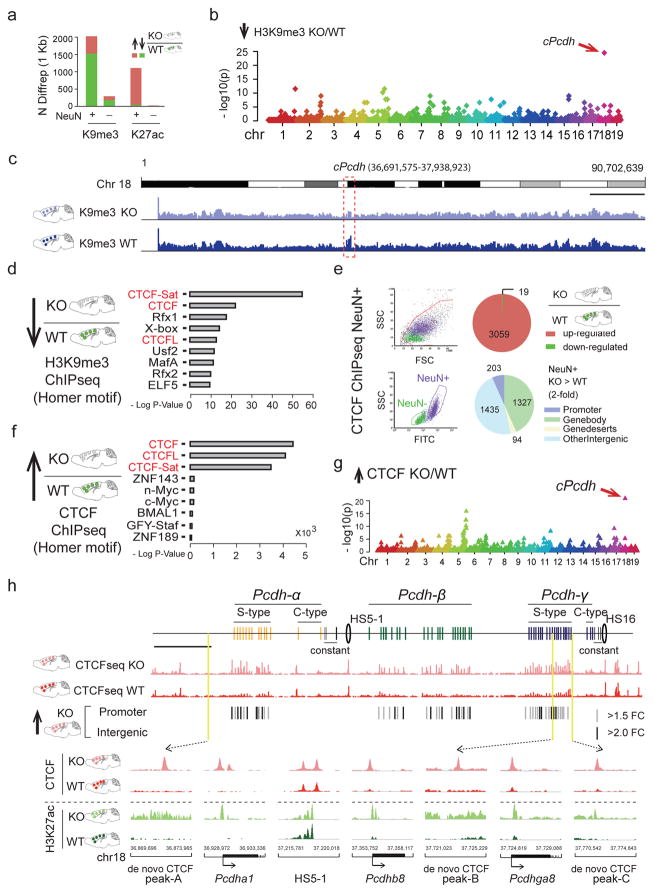
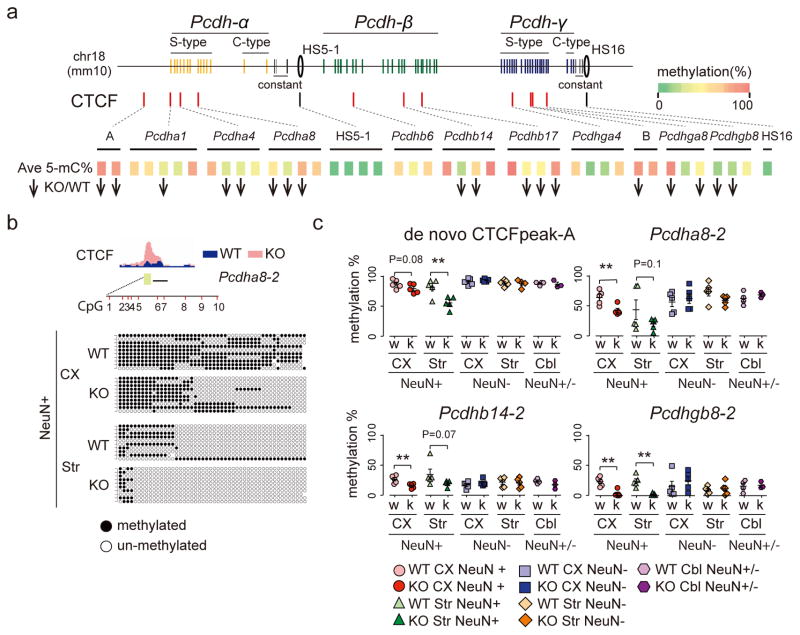
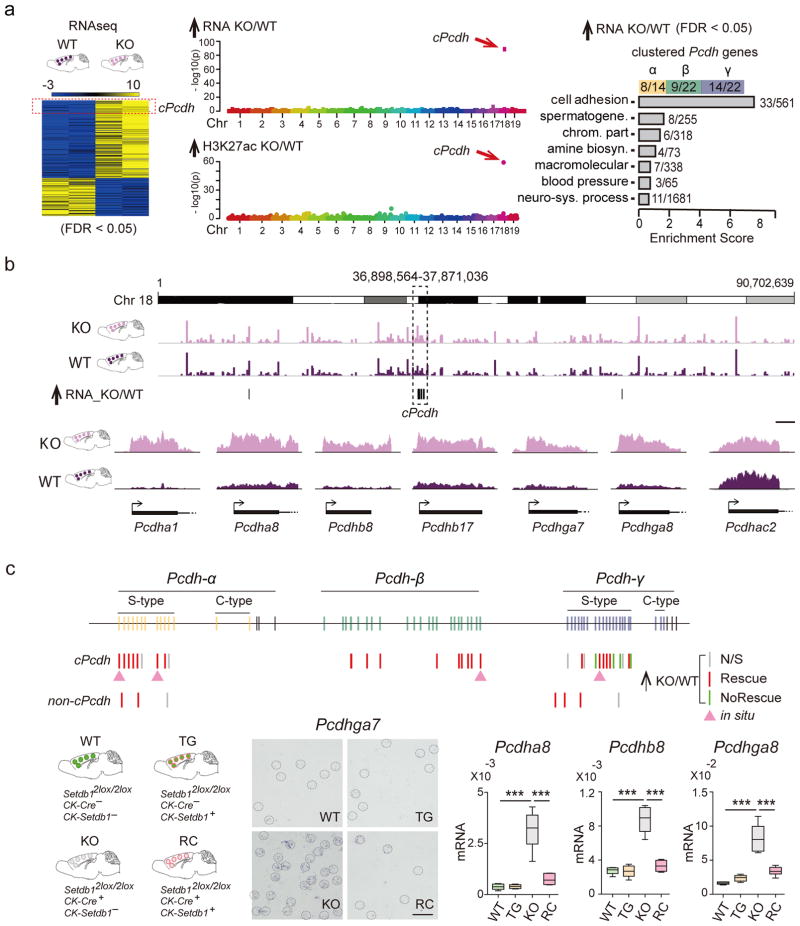
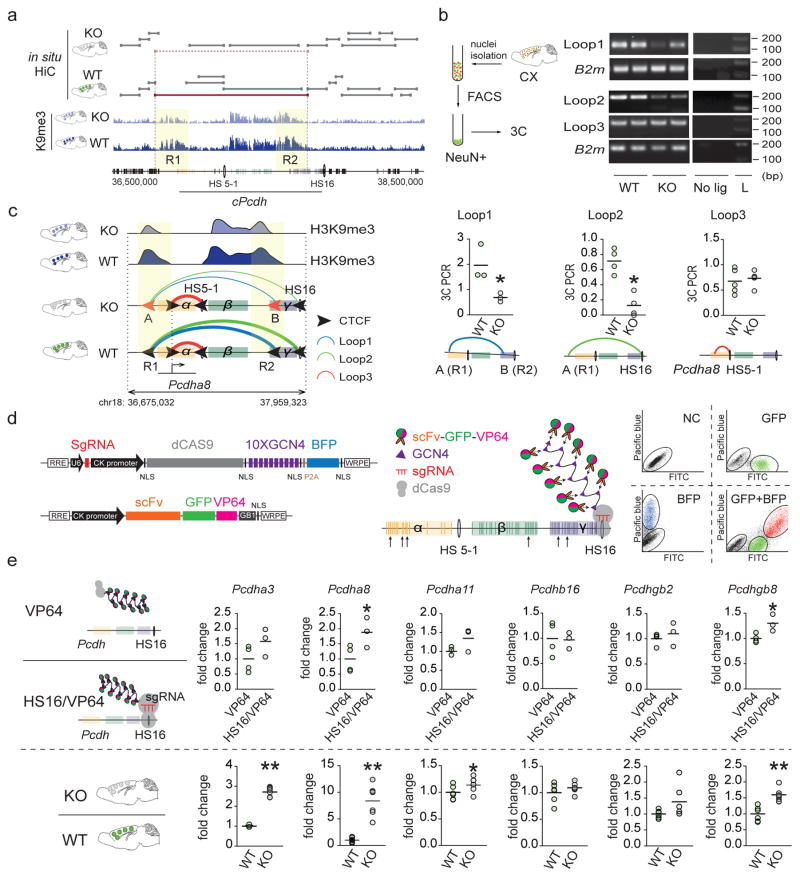
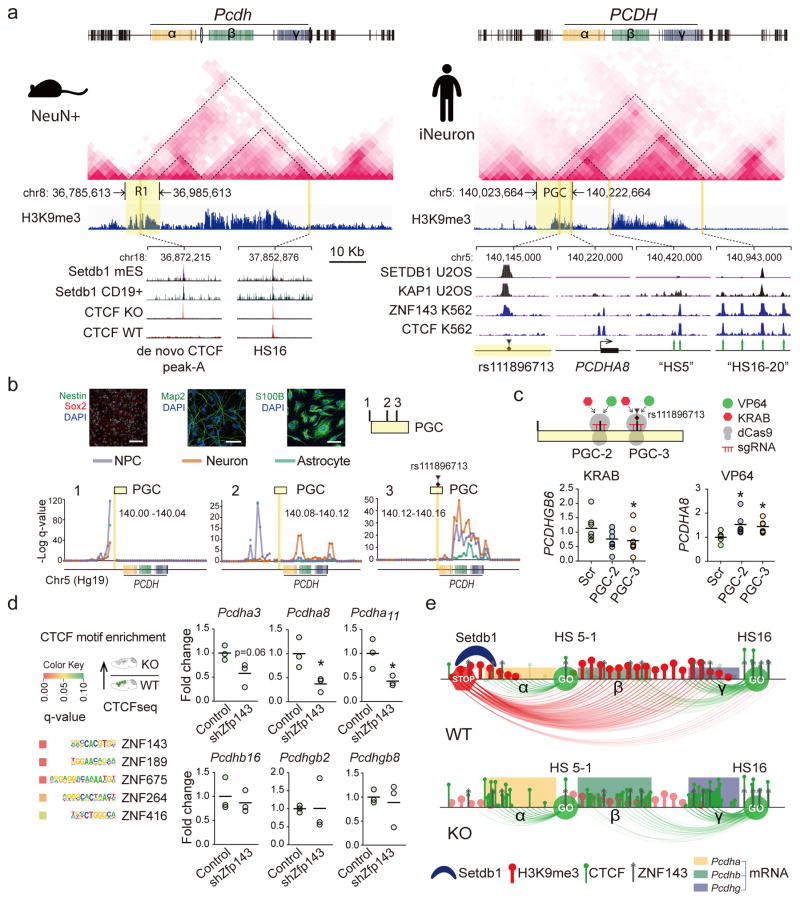
We report locus-specific disintegration of megabase-scale chromosomal conformations in brain after neuronal ablation of Setdb1 (also known as Kmt1e; encodes a histone H3 lysine 9 methyltransferase), including a large topologically associated 1.2-Mb domain conserved in humans and mice that encompasses >70 genes at the clustered protocadherin locus (hereafter referred to as cPcdh). The cPcdh topologically associated domain (TADcPcdh) in neurons from mutant mice showed abnormal accumulation of the transcriptional regulator and three-dimensional (3D) genome organizer CTCF at cryptic binding sites, in conjunction with DNA cytosine hypomethylation, histone hyperacetylation and upregulated expression. Genes encoding stochastically expressed protocadherins were transcribed by increased numbers of cortical neurons, indicating relaxation of single-cell constraint. SETDB1-dependent loop formations bypassed 0.2-1 Mb of linear genome and radiated from the TADcPcdh fringes toward cis-regulatory sequences within the cPcdh locus, counterbalanced shorter-range facilitative promoter-enhancer contacts and carried loop-bound polymorphisms that were associated with genetic risk for schizophrenia. We show that the SETDB1 repressor complex, which involves multiple KRAB zinc finger proteins, shields neuronal genomes from excess CTCF binding and is critically required for structural maintenance of TADcPcdh.
Conflict of interest statement
Figures
References
-
- Merkenschlager M, Nora EP. CTCF and Cohesin in Genome Folding and Transcriptional Gene Regulation. Annu Rev Genomics Hum Genet. 2016;17:17–43. - PubMed
MeSH terms
Substances
Grants and funding
- R01 AG050986/AG/NIA NIH HHS/United States
- T32 AG049688/AG/NIA NIH HHS/United States
- U01 AG046170/AG/NIA NIH HHS/United States
- S10 OD018522/OD/NIH HHS/United States
- R01 MH109677/MH/NIMH NIH HHS/United States
- F30 MH113330/MH/NIMH NIH HHS/United States
- R01 MH106056/MH/NIMH NIH HHS/United States
- R21 MH106919/MH/NIMH NIH HHS/United States
- T32 MH087004/MH/NIMH NIH HHS/United States
- R01 MH101454/MH/NIMH NIH HHS/United States
- I01 BX002395/BX/BLRD VA/United States
- P50 AG005138/AG/NIA NIH HHS/United States
- P50 MH096890/MH/NIMH NIH HHS/United States
- R01 NS091574/NS/NINDS NIH HHS/United States
- R01 EY024918/EY/NEI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
