

Disruption of a horizontally transferred phytoene desaturase abolishes carotenoid accumulation and diapause in Tetranychus urticae
- PMID: 28674017
- PMCID: PMC5530703
- DOI: 10.1073/pnas.1706865114
Disruption of a horizontally transferred phytoene desaturase abolishes carotenoid accumulation and diapause in Tetranychus urticae
Abstract
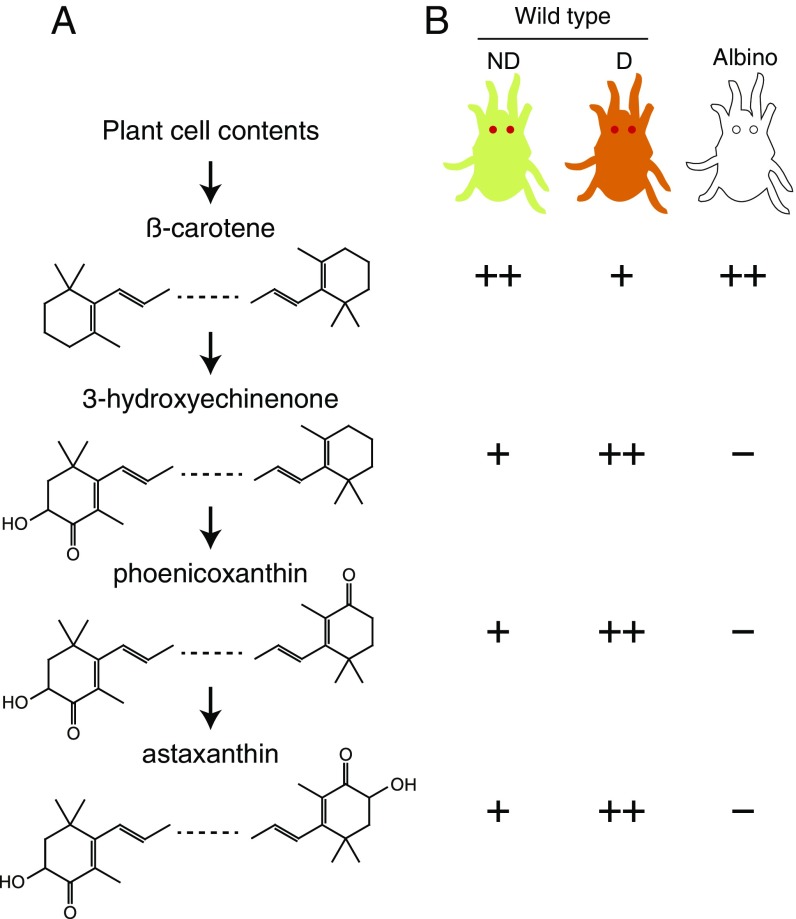
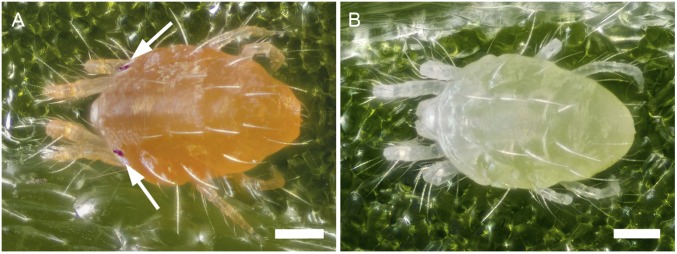
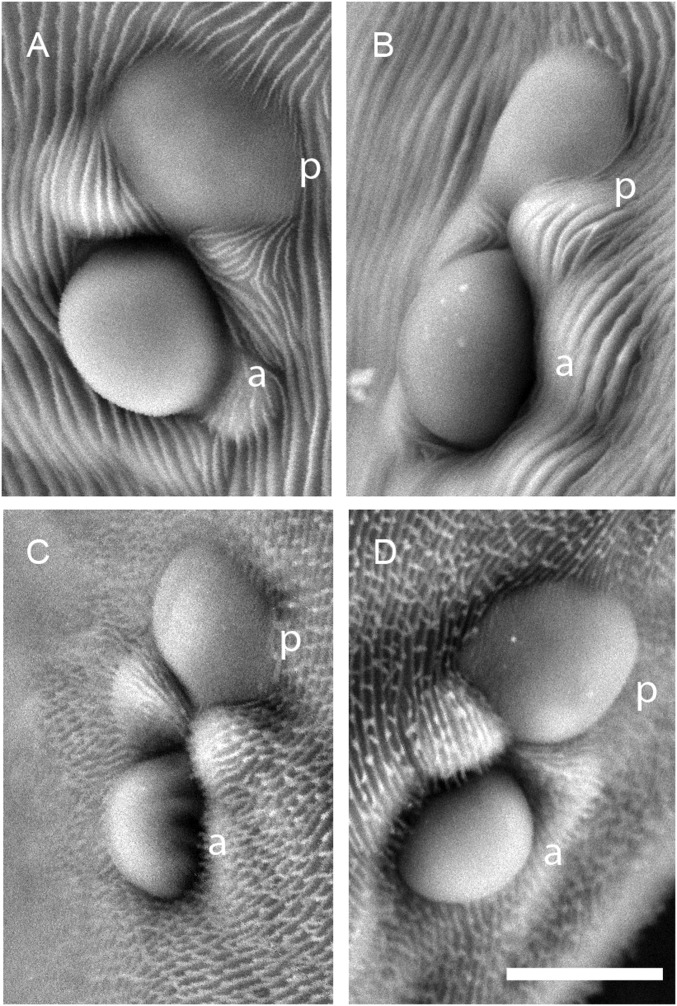
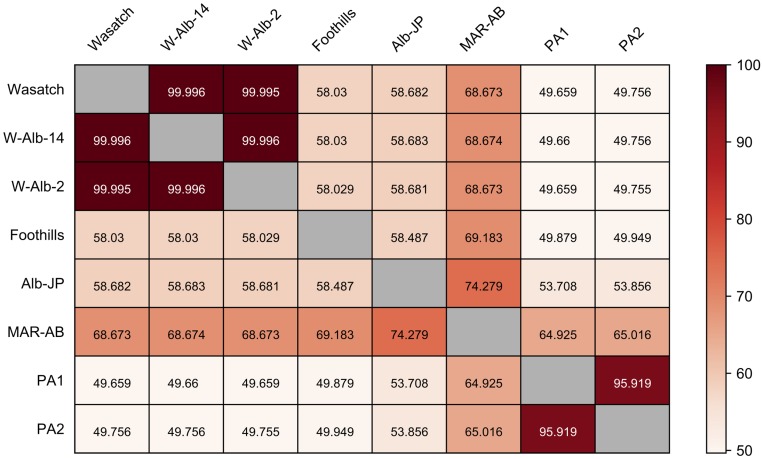
Carotenoids underlie many of the vibrant yellow, orange, and red colors in animals, and are involved in processes ranging from vision to protection from stresses. Most animals acquire carotenoids from their diets because de novo synthesis of carotenoids is primarily limited to plants and some bacteria and fungi. Recently, sequencing projects in aphids and adelgids, spider mites, and gall midges identified genes with homology to fungal sequences encoding de novo carotenoid biosynthetic proteins like phytoene desaturase. The finding of horizontal gene transfers of carotenoid biosynthetic genes to three arthropod lineages was unprecedented; however, the relevance of the transfers for the arthropods that acquired them has remained largely speculative, which is especially true for spider mites that feed on plant cell contents, a known source of carotenoids. Pigmentation in spider mites results solely from carotenoids. Using a combination of genetic approaches, we show that mutations in a single horizontally transferred phytoene desaturase result in complete albinism in the two-spotted spider mite, Tetranychus urticae, as well as in the citrus red mite, Panonychus citri Further, we show that phytoene desaturase activity is essential for photoperiodic induction of diapause in an overwintering strain of T. urticae, consistent with a role for this enzyme in provisioning provitamin A carotenoids required for light perception. Carotenoid biosynthetic genes of fungal origin have therefore enabled some mites to forgo dietary carotenoids, with endogenous synthesis underlying their intense pigmentation and ability to enter diapause, a key to the global distribution of major spider mite pests of agriculture.
Keywords: bulked segregant analysis; horizontal gene transfer; spider mites; xanthophylls; β-carotene.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Horizontally transferred fungal carotenoid genes in the two-spotted spider mite Tetranychus urticae.Biol Lett. 2012 Apr 23;8(2):253-7. doi: 10.1098/rsbl.2011.0704. Epub 2011 Sep 14. Biol Lett. 2012. PMID: 21920958 Free PMC article.
-
Molecular basis governing diapause and pigmentation in the hawthorn spider mite, Amphitetranychus viennensis.BMC Biol. 2025 Jun 3;23(1):152. doi: 10.1186/s12915-025-02258-z. BMC Biol. 2025. PMID: 40462063 Free PMC article.
-
Carotenoids in unexpected places: gall midges, lateral gene transfer, and carotenoid biosynthesis in animals.Mol Phylogenet Evol. 2013 Aug;68(2):221-8. doi: 10.1016/j.ympev.2013.03.012. Epub 2013 Mar 27. Mol Phylogenet Evol. 2013. PMID: 23542649
-
Physiological and molecular mechanisms underlying photoperiodism in the spider mite: comparisons with insects.J Comp Physiol B. 2016 Dec;186(8):969-984. doi: 10.1007/s00360-016-1018-9. Epub 2016 Jul 16. J Comp Physiol B. 2016. PMID: 27424162 Review.
-
Spider-mite problems and control in Taiwan.Exp Appl Acarol. 2000;24(5-6):453-62. doi: 10.1023/a:1006443619632. Exp Appl Acarol. 2000. PMID: 11156169 Review.
Cited by
-
Microbial platforms to produce commercially vital carotenoids at industrial scale: an updated review of critical issues.J Ind Microbiol Biotechnol. 2019 May;46(5):657-674. doi: 10.1007/s10295-018-2104-7. Epub 2018 Nov 10. J Ind Microbiol Biotechnol. 2019. PMID: 30415292 Review.
-
Carotenoid modifying enzymes in metazoans.Methods Enzymol. 2022;674:405-445. doi: 10.1016/bs.mie.2022.05.005. Epub 2022 Jun 22. Methods Enzymol. 2022. PMID: 36008015 Free PMC article.
-
Astaxanthin: Past, Present, and Future.Mar Drugs. 2023 Sep 28;21(10):514. doi: 10.3390/md21100514. Mar Drugs. 2023. PMID: 37888449 Free PMC article. Review.
-
Intraspecific diversity in the mechanisms underlying abamectin resistance in a cosmopolitan pest.Evol Appl. 2023 Mar 25;16(4):863-879. doi: 10.1111/eva.13542. eCollection 2023 Apr. Evol Appl. 2023. PMID: 37124092 Free PMC article.
-
Carotenoids: Distribution, Function in Nature, and Analysis Using LC-Photodiode Array Detector (DAD)-MS and MS/MS System.Mass Spectrom (Tokyo). 2023;12(1):A0133. doi: 10.5702/massspectrometry.A0133. Epub 2023 Nov 1. Mass Spectrom (Tokyo). 2023. PMID: 37937116 Free PMC article.
References
-
- Britton G. Structure and properties of carotenoids in relation to function. FASEB J. 1995;9:1551–1558. - PubMed
-
- Heath JJ, Cipollini DF, Stireman JO. The role of carotenoids and their derivatives in mediating interactions between insects and their environment. Arthropod-Plant Interact. 2013;7:1–20.
-
- Bendich A, Olson JA. Biological actions of carotenoids. FASEB J. 1989;3:1927–1932. - PubMed
-
- Sies H, Stahl W. Vitamins E and C, beta-carotene, and other carotenoids as antioxidants. Am J Clin Nutr. 1995;62(6) Suppl:1315S–1321S. - PubMed
-
- Stahl W, Ale-Agha N, Polidori MC. Non-antioxidant properties of carotenoids. Biol Chem. 2002;383:553–558. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
