

Distinct Correlation Structure Supporting a Rate-Code for Sound Localization in the Owl's Auditory Forebrain
- PMID: 28674698
- PMCID: PMC5492684
- DOI: 10.1523/ENEURO.0144-17.2017
Distinct Correlation Structure Supporting a Rate-Code for Sound Localization in the Owl's Auditory Forebrain
Abstract
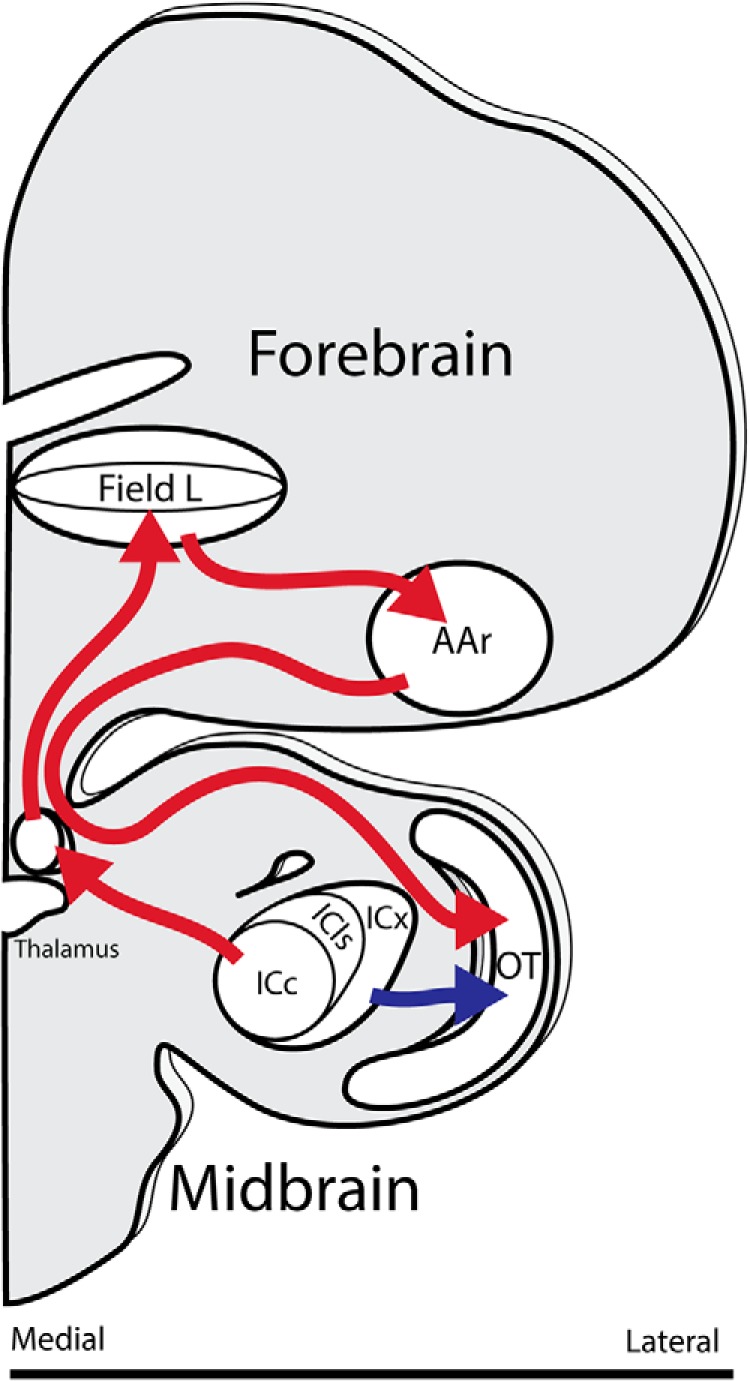
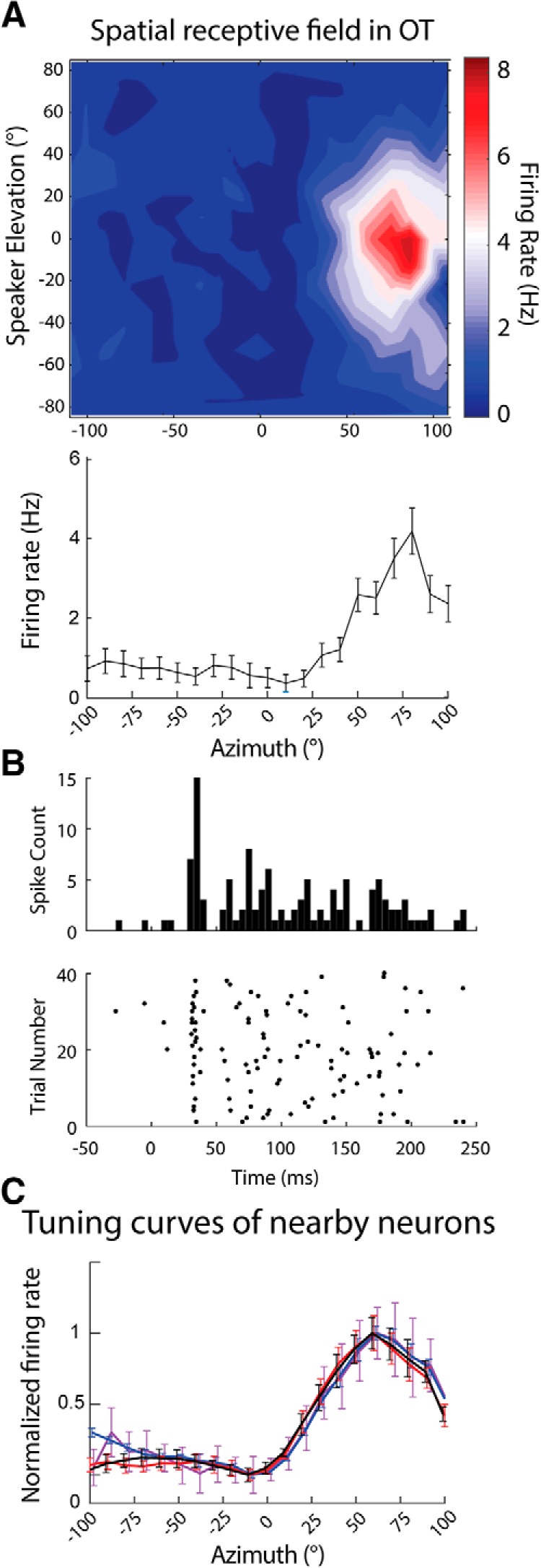
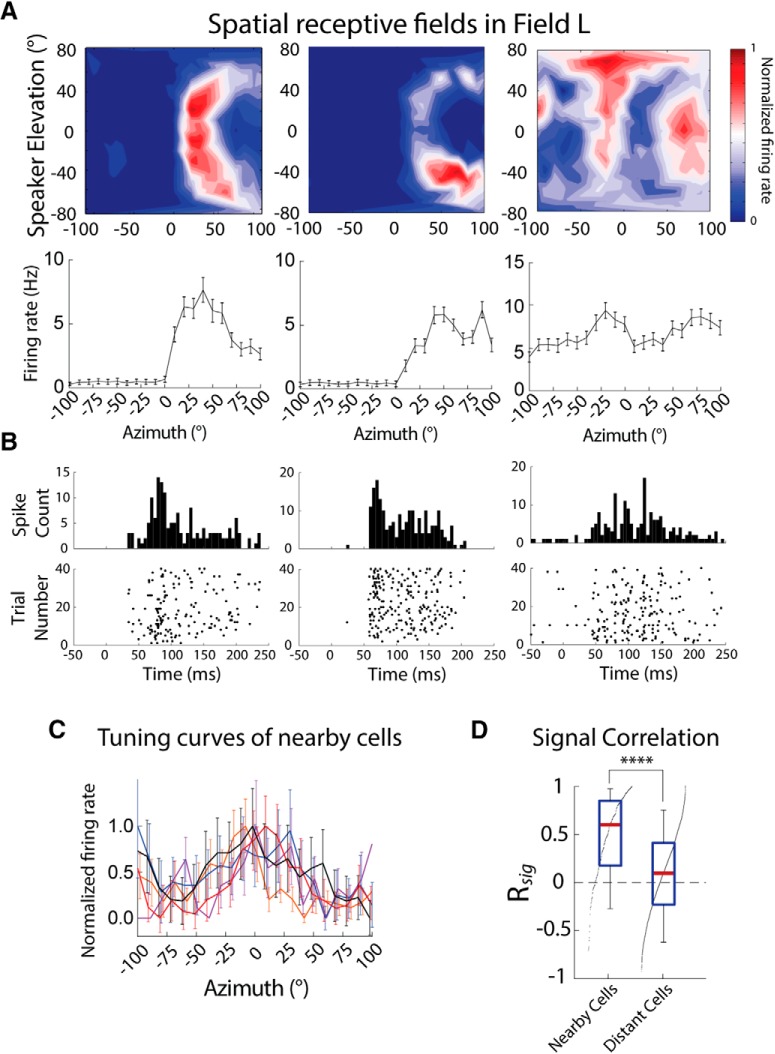
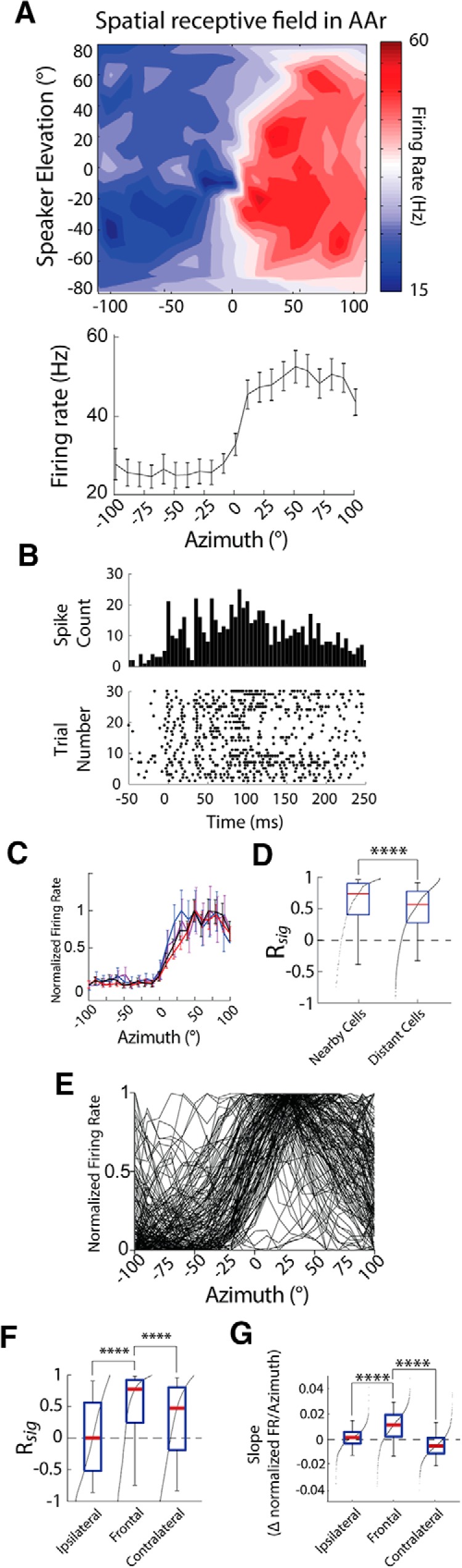
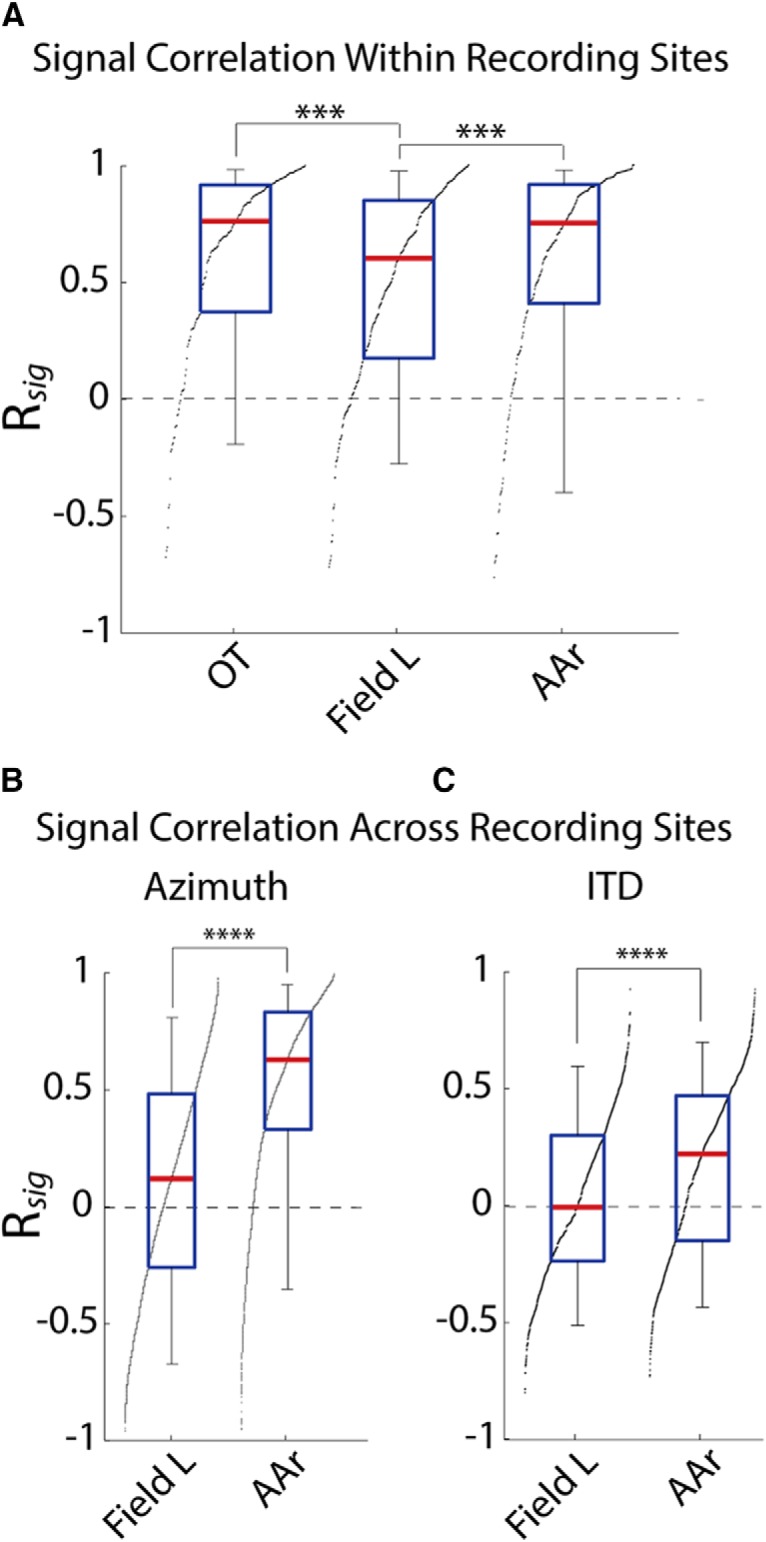
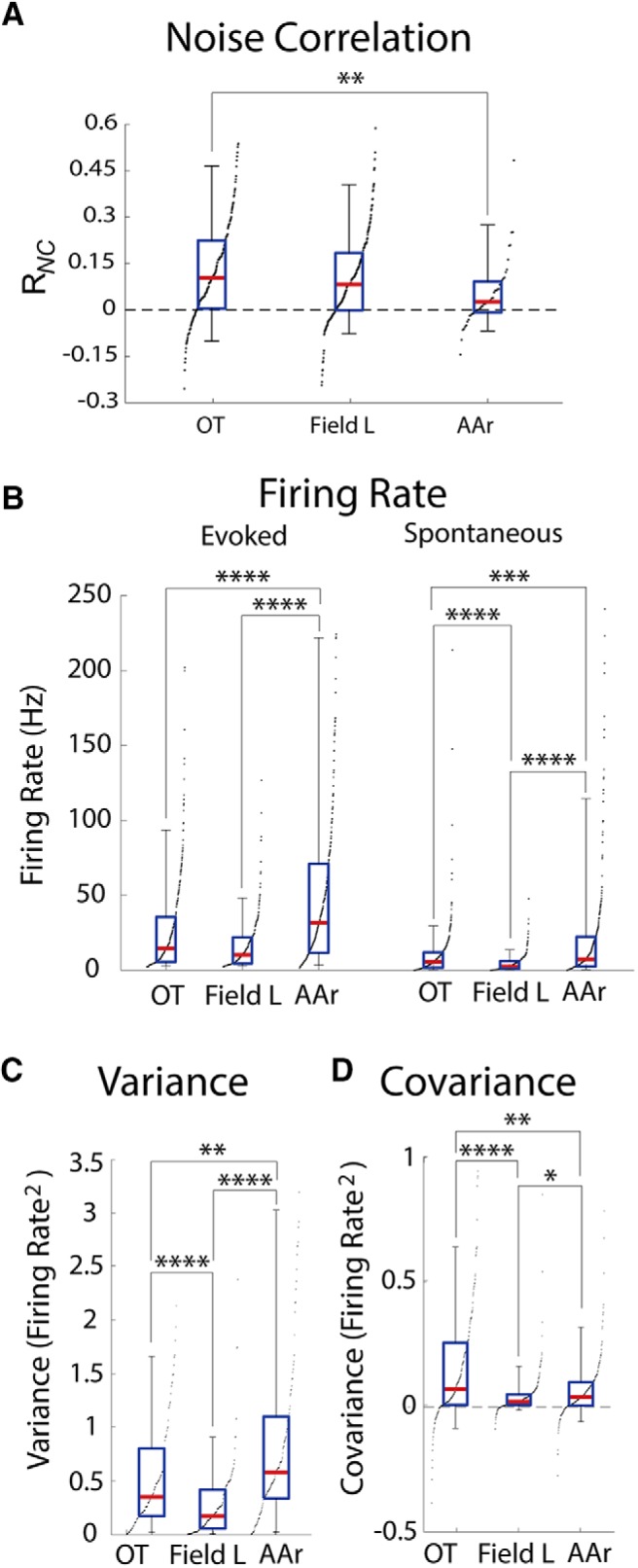
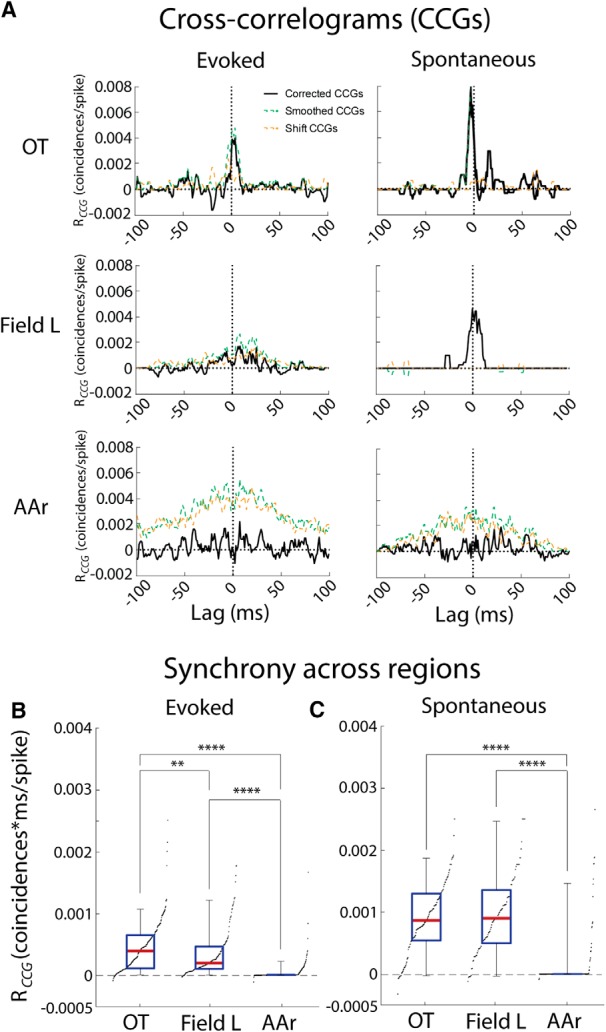
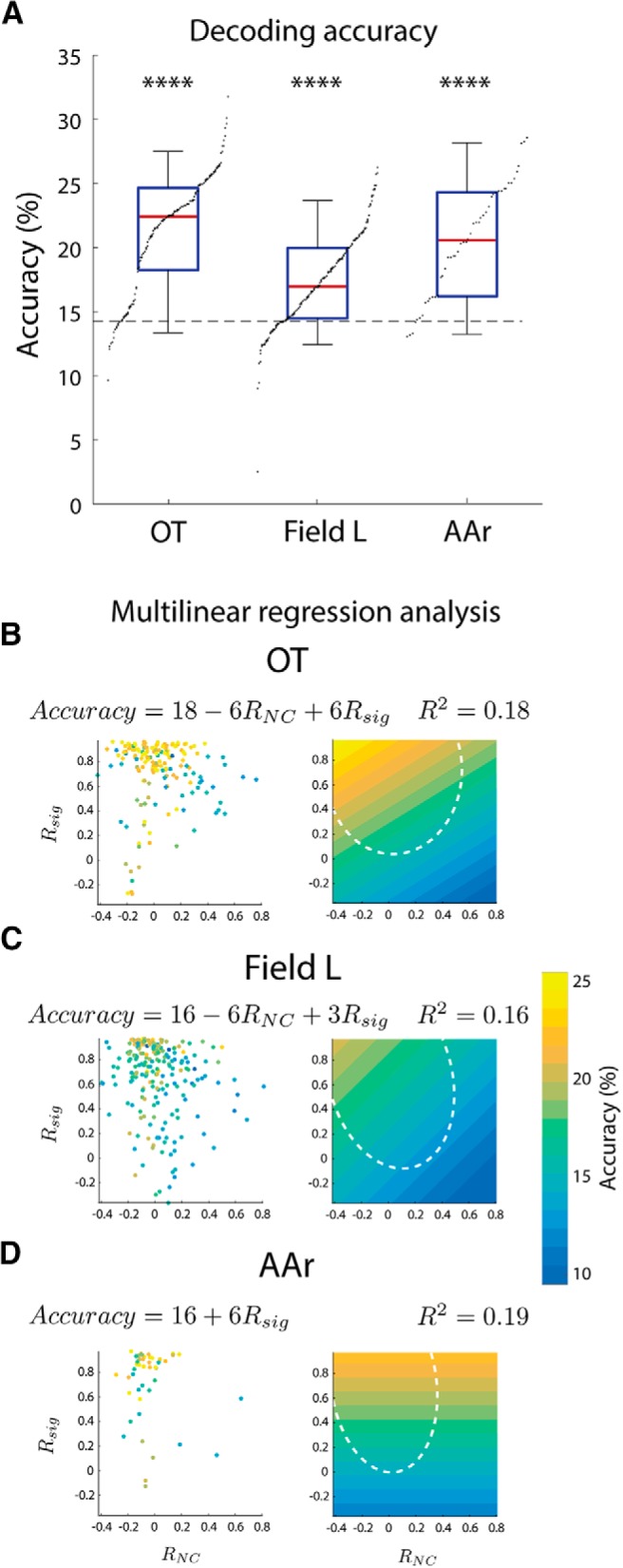
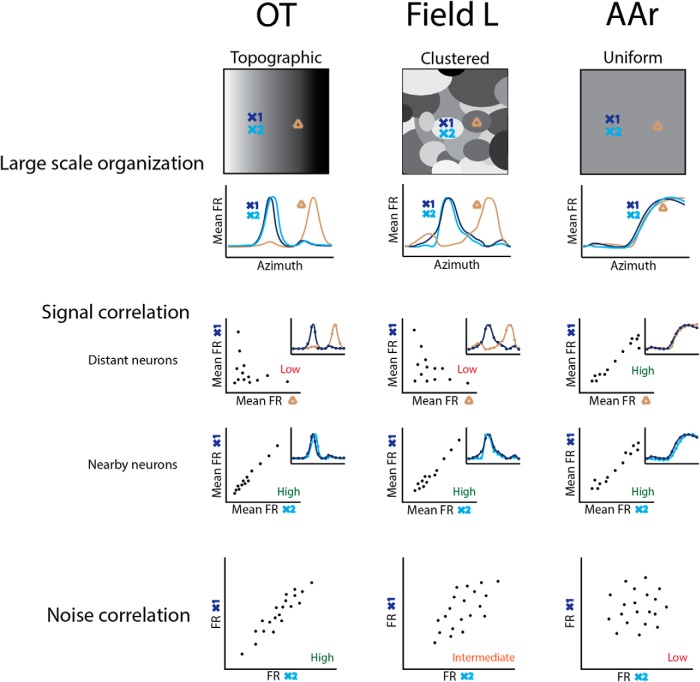
While a topographic map of auditory space exists in the vertebrate midbrain, it is absent in the forebrain. Yet, both brain regions are implicated in sound localization. The heterogeneous spatial tuning of adjacent sites in the forebrain compared to the midbrain reflects different underlying circuitries, which is expected to affect the correlation structure, i.e., signal (similarity of tuning) and noise (trial-by-trial variability) correlations. Recent studies have drawn attention to the impact of response correlations on the information readout from a neural population. We thus analyzed the correlation structure in midbrain and forebrain regions of the barn owl's auditory system. Tetrodes were used to record in the midbrain and two forebrain regions, Field L and the downstream auditory arcopallium (AAr), in anesthetized owls. Nearby neurons in the midbrain showed high signal and noise correlations (R NC s), consistent with shared inputs. As previously reported, Field L was arranged in random clusters of similarly tuned neurons. Interestingly, AAr neurons displayed homogeneous monotonic azimuth tuning, while response variability of nearby neurons was significantly less correlated than the midbrain. Using a decoding approach, we demonstrate that low R NC in AAr restricts the potentially detrimental effect it can have on information, assuming a rate code proposed for mammalian sound localization. This study harnesses the power of correlation structure analysis to investigate the coding of auditory space. Our findings demonstrate distinct correlation structures in the auditory midbrain and forebrain, which would be beneficial for a rate-code framework for sound localization in the nontopographic forebrain representation of auditory space.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials