

Trace Conditioning in Drosophila Induces Associative Plasticity in Mushroom Body Kenyon Cells and Dopaminergic Neurons
- PMID: 28676744
- PMCID: PMC5476701
- DOI: 10.3389/fncir.2017.00042
Trace Conditioning in Drosophila Induces Associative Plasticity in Mushroom Body Kenyon Cells and Dopaminergic Neurons
Abstract
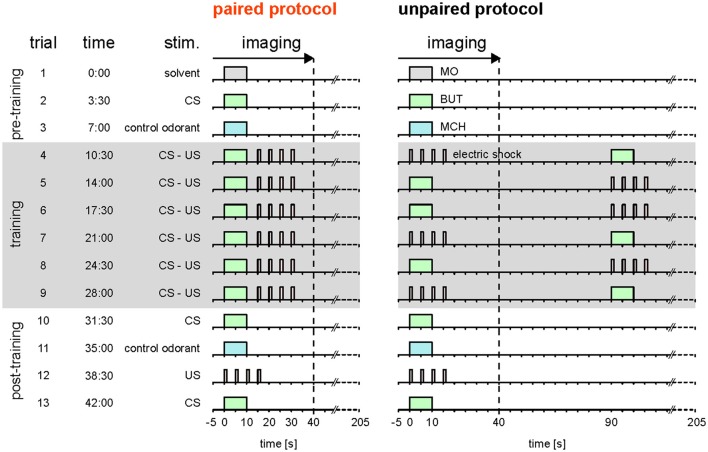
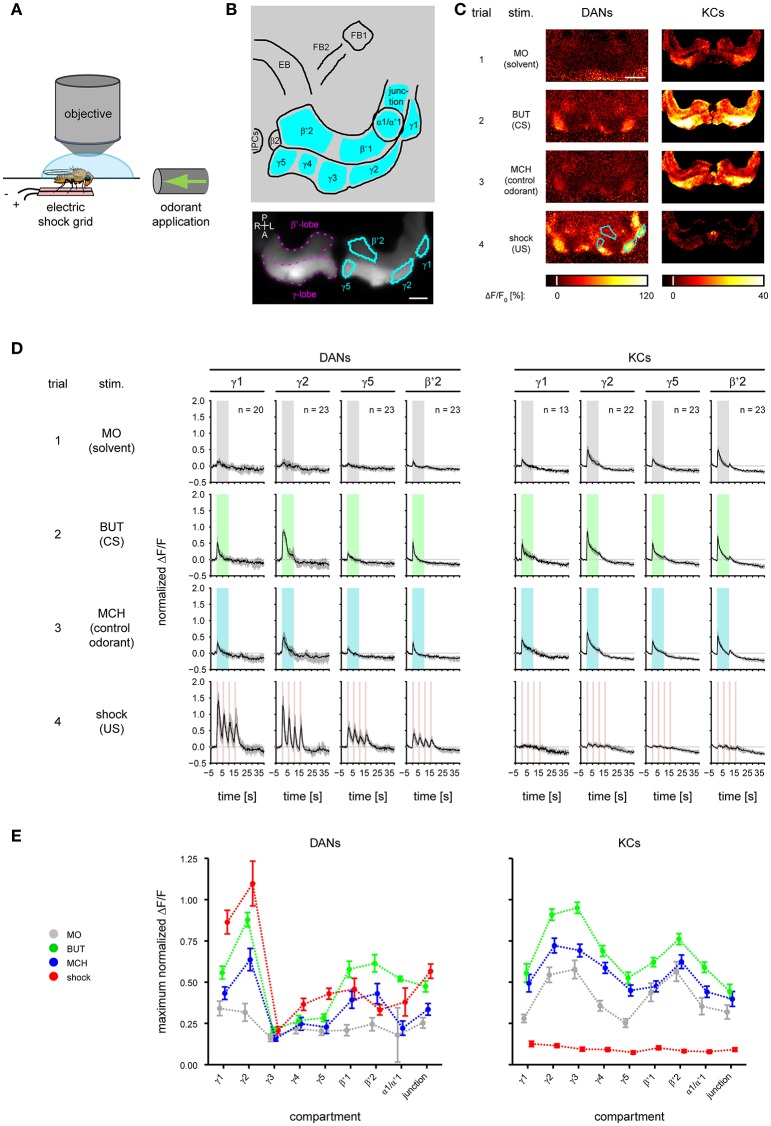
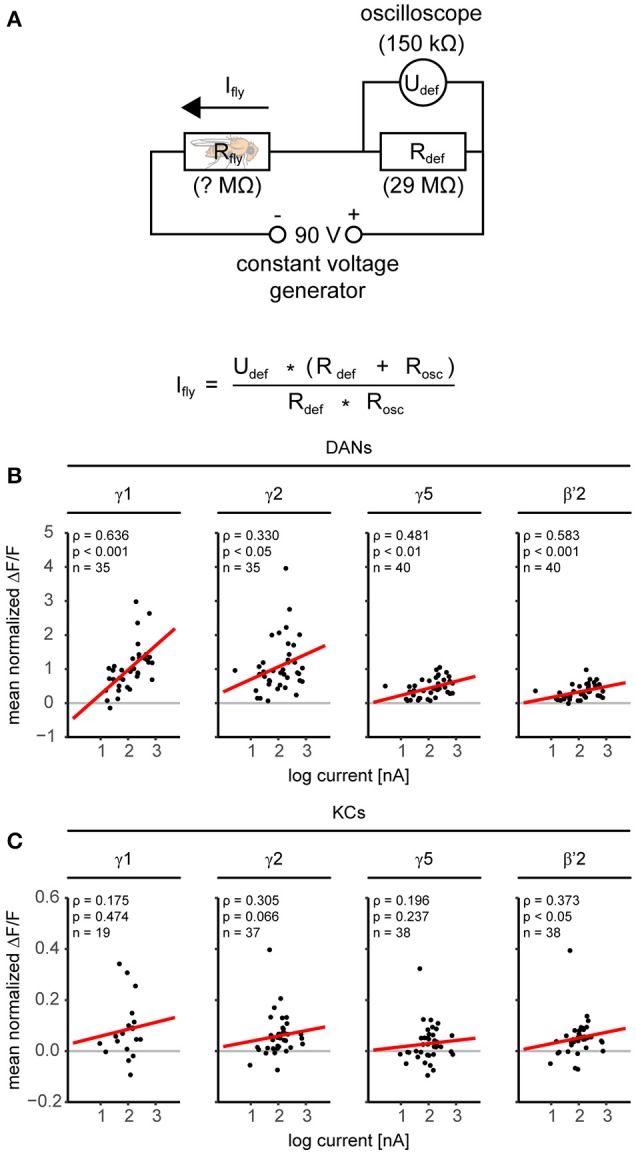
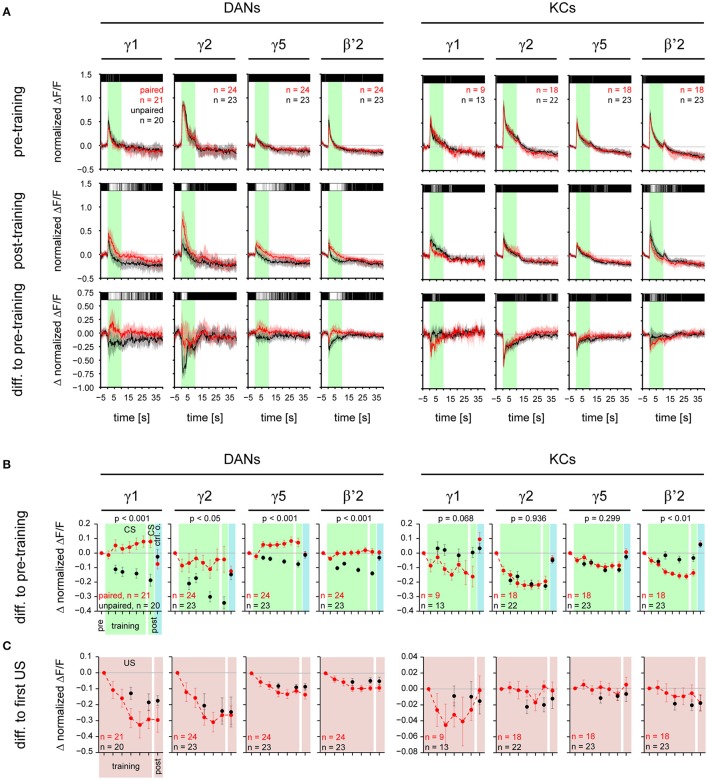
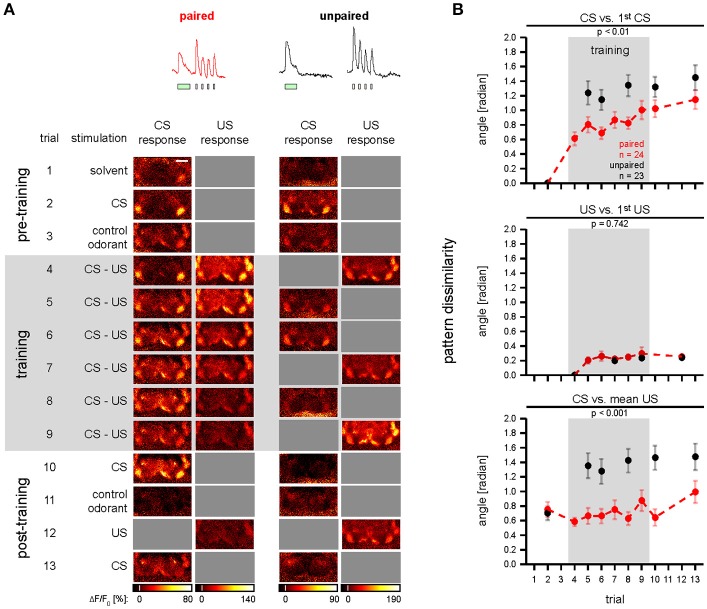
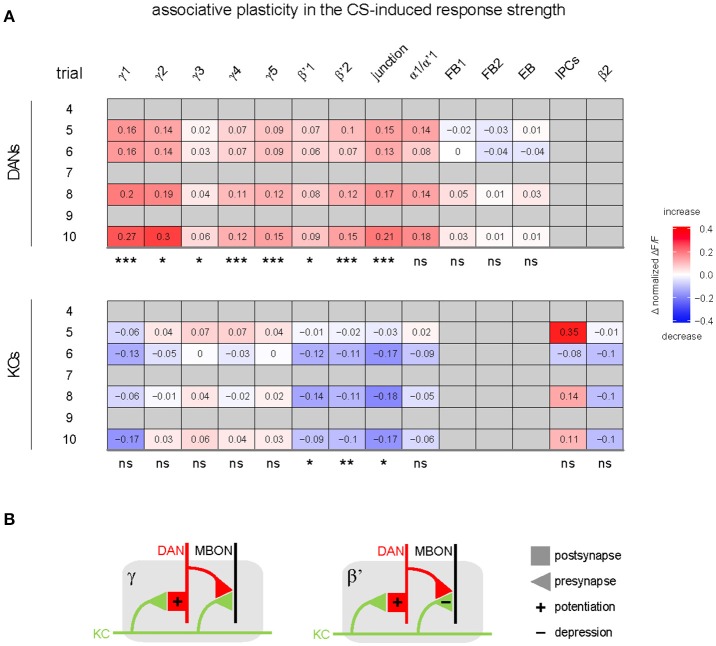
Dopaminergic neurons (DANs) signal punishment and reward during associative learning. In mammals, DANs show associative plasticity that correlates with the discrepancy between predicted and actual reinforcement (prediction error) during classical conditioning. Also in insects, such as Drosophila, DANs show associative plasticity that is, however, less understood. Here, we study associative plasticity in DANs and their synaptic partners, the Kenyon cells (KCs) in the mushroom bodies (MBs), while training Drosophila to associate an odorant with a temporally separated electric shock (trace conditioning). In most MB compartments DANs strengthened their responses to the conditioned odorant relative to untrained animals. This response plasticity preserved the initial degree of similarity between the odorant- and the shock-induced spatial response patterns, which decreased in untrained animals. Contrary to DANs, KCs (α'/β'-type) decreased their responses to the conditioned odorant relative to untrained animals. We found no evidence for prediction error coding by DANs during conditioning. Rather, our data supports the hypothesis that DAN plasticity encodes conditioning-induced changes in the odorant's predictive power.
Keywords: Drosophila; Kenyon cells; associative plasticity; calcium imaging; dopaminergic neurons; memory acquisition; mushroom body; trace conditioning.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous