

Karyological evidence of hybridogenesis in Greenlings (Teleostei: Hexagrammidae)
- PMID: 28678883
- PMCID: PMC5498075
- DOI: 10.1371/journal.pone.0180626
Karyological evidence of hybridogenesis in Greenlings (Teleostei: Hexagrammidae)
Abstract
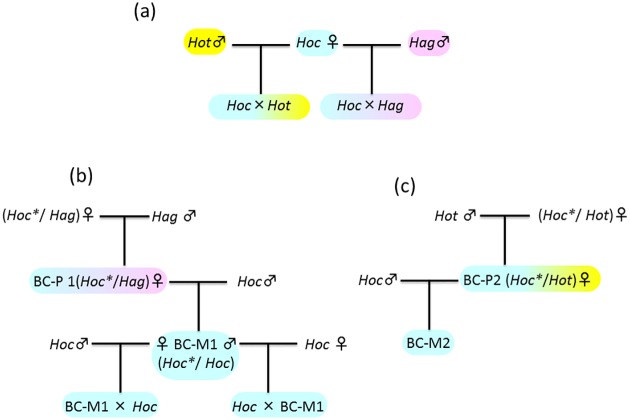
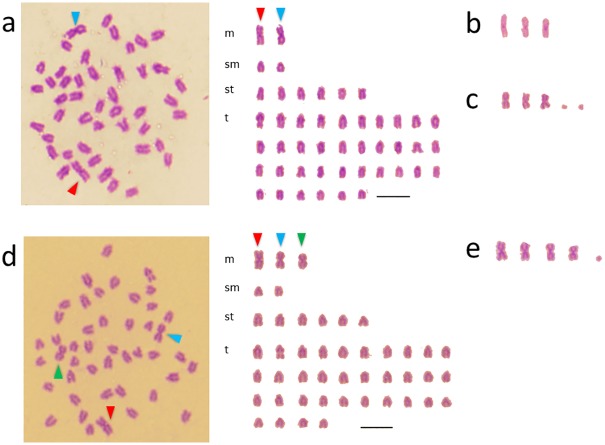
Two types of natural hybrids were discovered in populations of three Hexagrammos species (Teleostei: Hexagrammidae) distributed off the southern coast of Hokkaido in the North Pacific Ocean. Both hybrids reproduce by hybridogenesis, in which the maternal haploid genome is transmitted to offspring without recombination and the paternal haploid genome is eliminated during gametogenesis. While natural hybrids are unisexual and reproduce hemiclonally by backcrossing with the paternal species (BC-P), artificial F1-hybrids between the pure species produce recombinant gametes. Thus, despite having the same genome composition, the natural hybrids and the F1-hybrids are not genetically identical. Here, to clarify the differences between both hybrids, we examined the karyotypes of the three Hexagrammos species, their natural hybrids, the artificial F1-hybrids, and several backcrosses. Artificial F1-hybrids have karyotypes and chromosome numbers that are intermediate between those of the parental species. Conversely, the natural hybrids differed from F1-hybrids by having several large metacentric chromosomes and microchromosomes. Since the entire maternal haploid genome is inherited by the natural hybrids, maternal backcrosses (BC-M) between natural hybrids and males of the maternal species (H. octogrammus; Hoc) have a hemiclonal Hoc genome with large chromosomes from the mother and a normal Hoc genome from the father. However, the large chromosomes disappear in offspring of BC-M, probably due to fissuring during gametogenesis. Similarly, microsatellite DNA analysis revealed that chromosomes of BC-M undergo recombination. These findings suggest that genetic factors associated with hemiclonal reproduction may be located on the large metacentric chromosomes of natural hybrids.
Conflict of interest statement
Figures




Similar articles
-
A robust system of hybridogenesis that increases genetic variability and promotes evolutionary succession in greenlings (Teleostei: Hexagrammidae, genus Hexagrammos): Regeneration of a new hemiclonal lineage.PLoS One. 2024 Jun 3;19(6):e0304772. doi: 10.1371/journal.pone.0304772. eCollection 2024. PLoS One. 2024. PMID: 38829848 Free PMC article.
-
Identification of hemiclonal reproduction in three species of Hexagrammos marine reef fishes.J Fish Biol. 2014 Aug;85(2):189-209. doi: 10.1111/jfb.12414. Epub 2014 Jun 5. J Fish Biol. 2014. PMID: 24903212
-
Origins of two hemiclonal hybrids among three Hexagrammos species (Teleostei: Hexagrammidae): genetic diversification through host switching.Ecol Evol. 2016 Sep 14;6(19):7126-7140. doi: 10.1002/ece3.2446. eCollection 2016 Oct. Ecol Evol. 2016. PMID: 28725387 Free PMC article.
-
Genomic constitution and atypical reproduction in polyploid and unisexual lineages of the Misgurnus loach, a teleost fish.Cytogenet Genome Res. 2013;140(2-4):226-40. doi: 10.1159/000353301. Epub 2013 Jul 9. Cytogenet Genome Res. 2013. PMID: 23899809 Review.
-
Identification of parental chromosomes involved in translocations BCR-ABL, t(9;22) and PML-RARA, t(15;17).Anticancer Res. 2008 Nov-Dec;28(6A):3573-8. Anticancer Res. 2008. PMID: 19189637 Review.
Cited by
-
Using asexual vertebrates to study genome evolution and animal physiology: Banded (Fundulus diaphanus) x Common Killifish (F. heteroclitus) hybrid lineages as a model system.Evol Appl. 2020 May 4;13(6):1214-1239. doi: 10.1111/eva.12975. eCollection 2020 Jul. Evol Appl. 2020. PMID: 32684956 Free PMC article.
-
Uniparental Genome Elimination in Australian Carp Gudgeons.Genome Biol Evol. 2021 Jun 8;13(6):evab030. doi: 10.1093/gbe/evab030. Genome Biol Evol. 2021. PMID: 33591327 Free PMC article.
-
Hybrid between Danio rerio female and Danio nigrofasciatus male produces aneuploid sperm with limited fertilization capacity.PLoS One. 2020 May 29;15(5):e0233885. doi: 10.1371/journal.pone.0233885. eCollection 2020. PLoS One. 2020. PMID: 32470029 Free PMC article.
-
A robust system of hybridogenesis that increases genetic variability and promotes evolutionary succession in greenlings (Teleostei: Hexagrammidae, genus Hexagrammos): Regeneration of a new hemiclonal lineage.PLoS One. 2024 Jun 3;19(6):e0304772. doi: 10.1371/journal.pone.0304772. eCollection 2024. PLoS One. 2024. PMID: 38829848 Free PMC article.
References
-
- Arnold ML. Natural hybridization and evolution. Oxford University Press; 1997.
-
- Coyne JA, Orr HA. Speciation. Sinauer Associates In: Massachusettus. 2004.
-
- Vrijenhoek RC, Dawley R, Cole CJ, Bogart JP. A list of the known unisexual vertebrates In: Dawley M, Bogart JP. Evolution and Ecology of Unisexual Vertebrates, New York State Museum; 1989. pp.19–23.
-
- Vrijenhoek RC. Animal clones and diversity. Bioscience. 1998; 48: 617–628.
-
- Burt A, Trivers R. Genes in Conflict: The Biology of Selfish Genetic Elements. Belknap Press of Harvard University Press; 2006.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
