

Basal hypersecretion of glucagon and insulin from palmitate-exposed human islets depends on FFAR1 but not decreased somatostatin secretion
- PMID: 28680093
- PMCID: PMC5498543
- DOI: 10.1038/s41598-017-04730-5
Basal hypersecretion of glucagon and insulin from palmitate-exposed human islets depends on FFAR1 but not decreased somatostatin secretion
Abstract
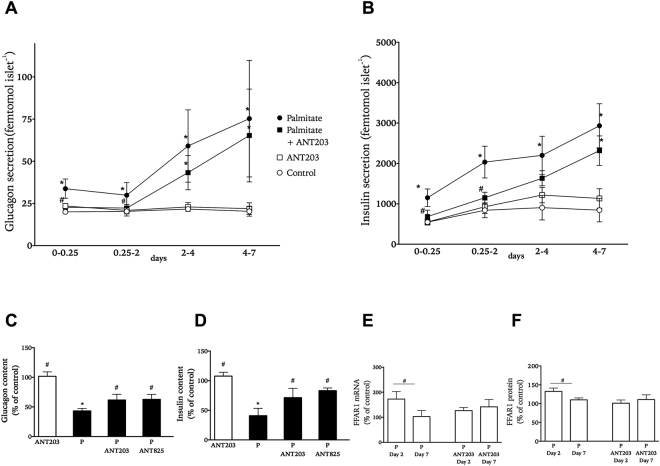
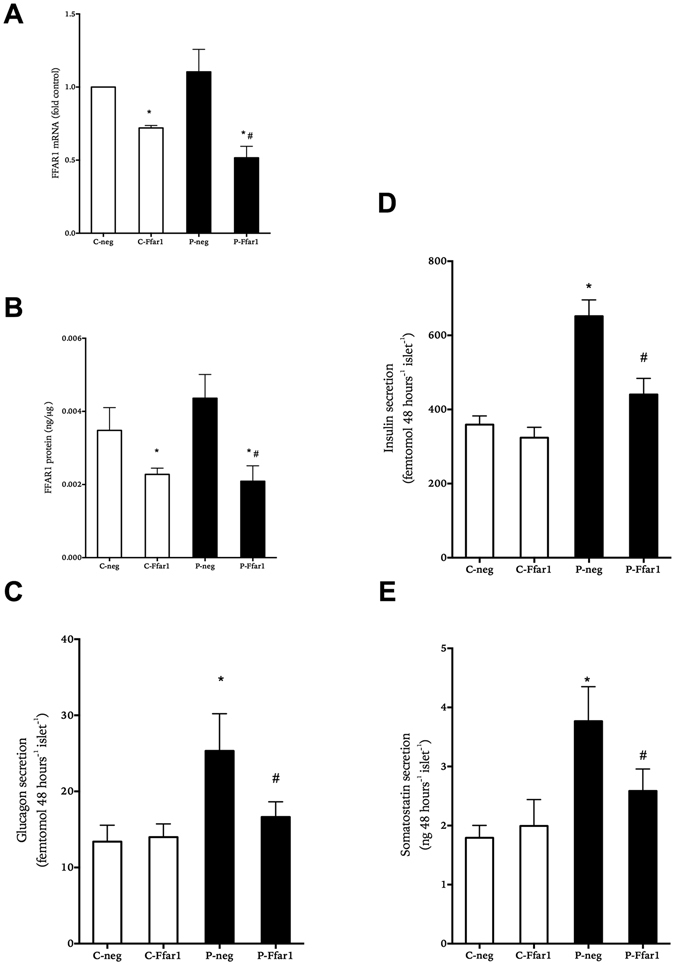
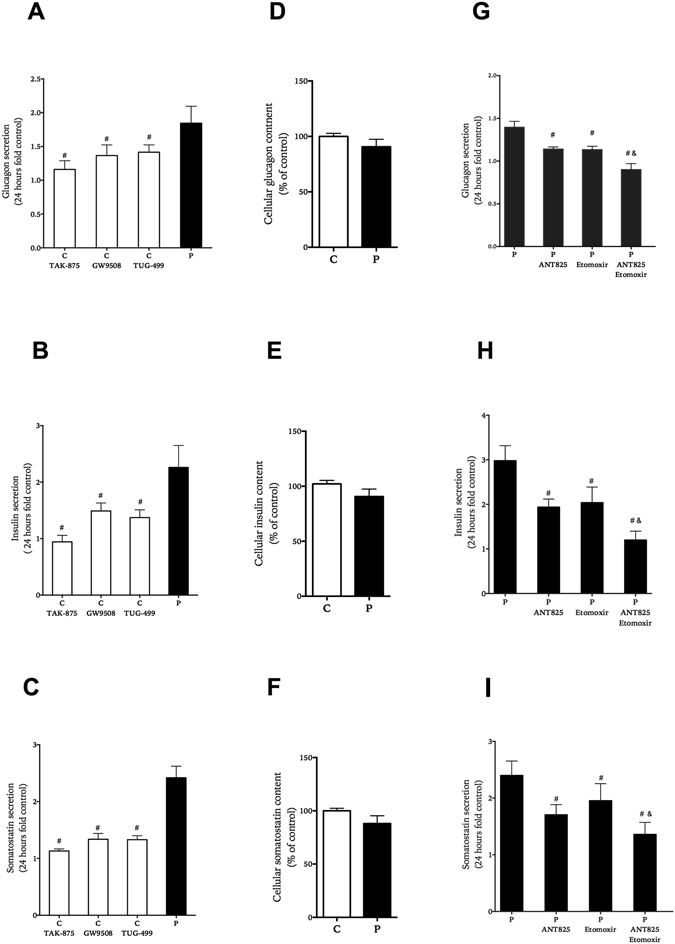
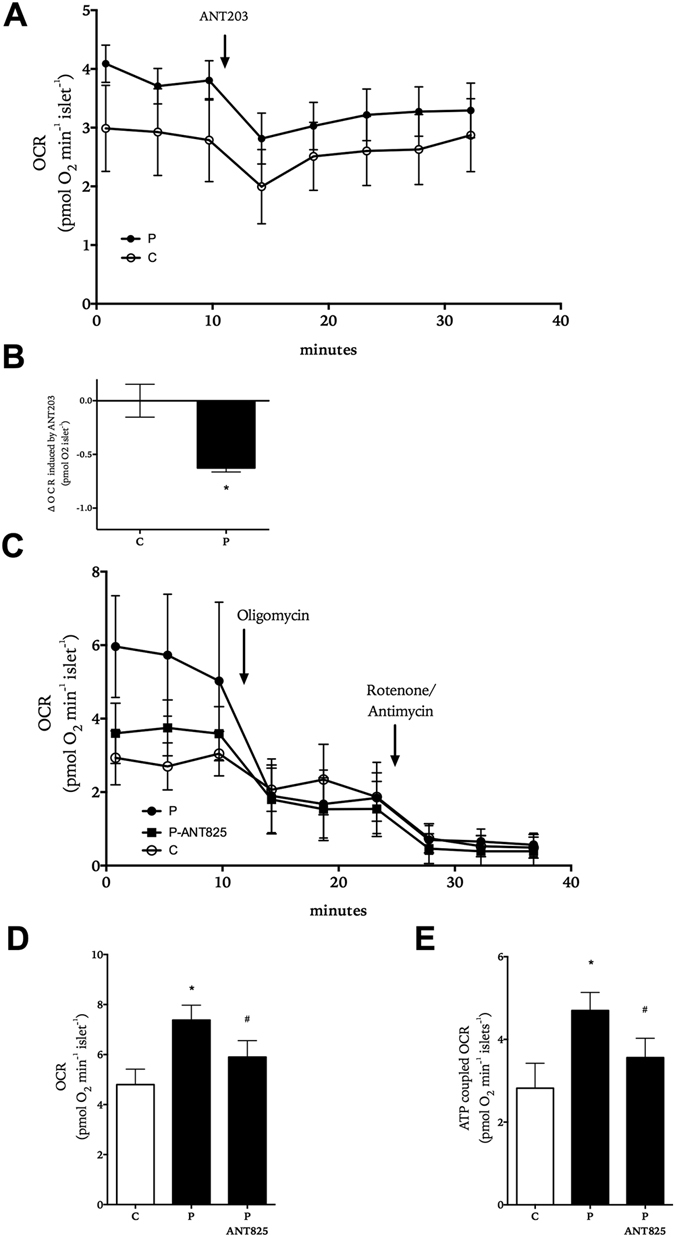
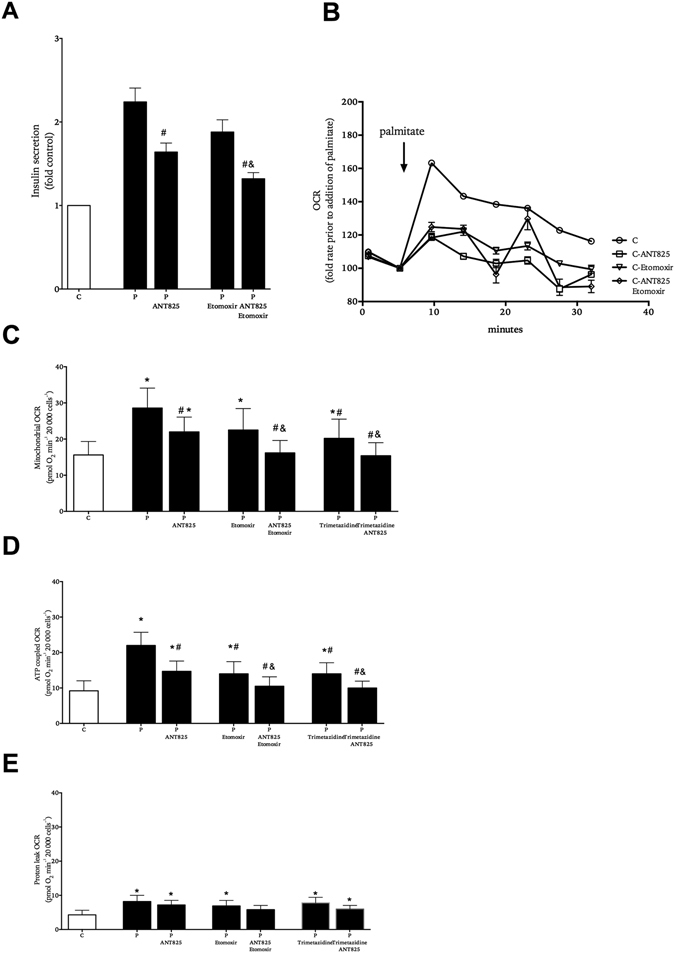
In obesity fasting levels of both glucagon and insulin are elevated. In these subjects fasting levels of the free fatty acid palmitate are raised. We have demonstrated that palmitate enhances glucose-stimulated insulin secretion from isolated human islets via free fatty acid receptor 1 (FFAR1/GPR40). Since FFAR1 is also present on glucagon-secreting alpha-cells, we hypothesized that palmitate simultaneously stimulates secretion of glucagon and insulin at fasting glucose concentrations. In addition, we hypothesized that concomitant hypersecretion of glucagon and insulin was also contributed by reduced somatostatin secretion. We found basal glucagon, insulin and somatostatin secretion and respiration from human islets, to be enhanced during palmitate treatment at normoglycemia. Secretion of all hormones and mitochondrial respiration were lowered when FFAR1 or fatty acid β-oxidation was inhibited. The findings were confirmed in the human beta-cell line EndoC-βH1. We conclude that fatty acids enhance both glucagon and insulin secretion at fasting glucose concentrations and that FFAR1 and enhanced mitochondrial metabolism but not lowered somatostatin secretion are crucial in this effect. The ability of chronically elevated palmitate levels to simultaneously increase basal secretion of glucagon and insulin positions elevated levels of fatty acids as potential triggering factors for the development of obesity and impaired glucose control.
Conflict of interest statement
H.K., E.S., H.M. and P.B. have nothing to disclose. S.O.G. and D.M.S. are employed by AstraZeneca.
Figures
References
-
- Erdmann J, et al. Disturbances of basal and postprandial insulin secretion and clearance in obese patients with type 2 diabetes mellitus. Hormone and metabolic research = Hormon- und Stoffwechselforschung = Hormones et metabolisme. 2012;44:60–69. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
