

Tumor-priming converts NK cells to memory-like NK cells
- PMID: 28680749
- PMCID: PMC5486172
- DOI: 10.1080/2162402X.2017.1317411
Tumor-priming converts NK cells to memory-like NK cells
Abstract
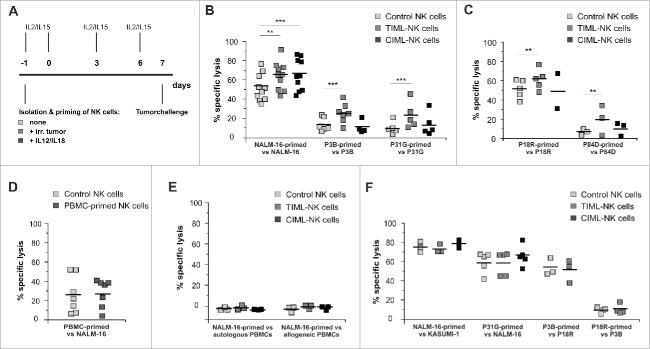
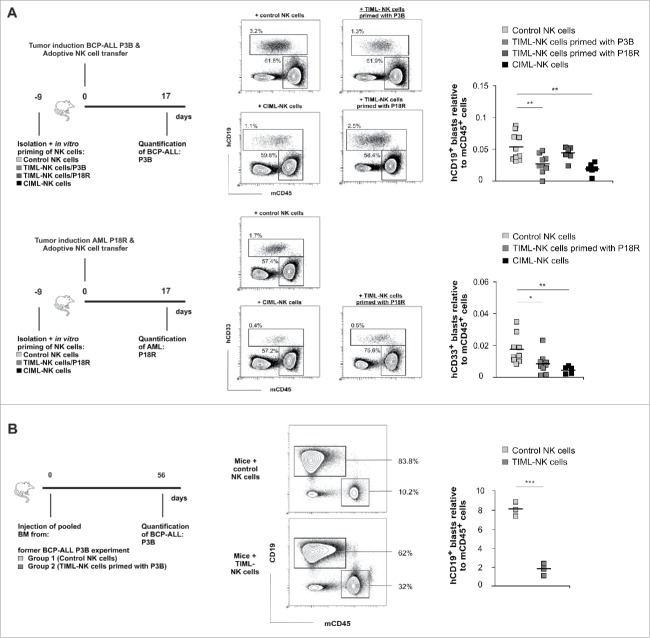
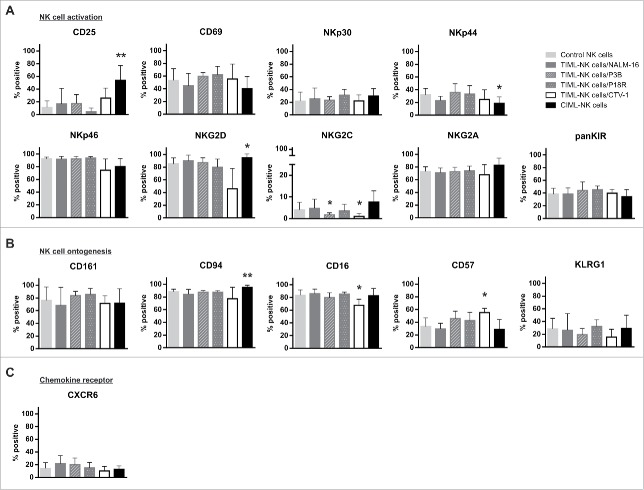
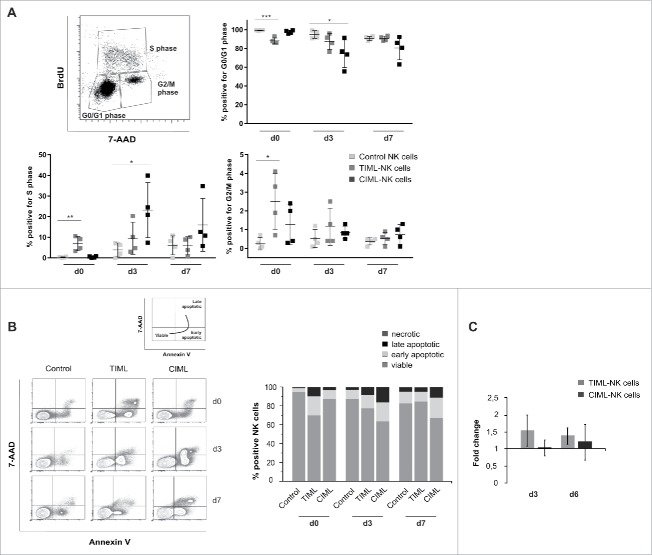
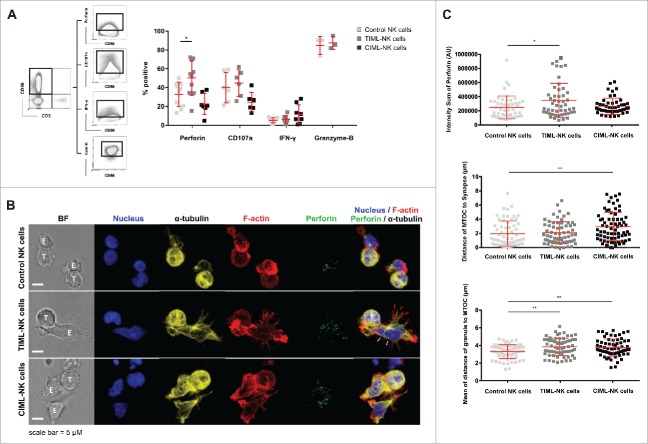
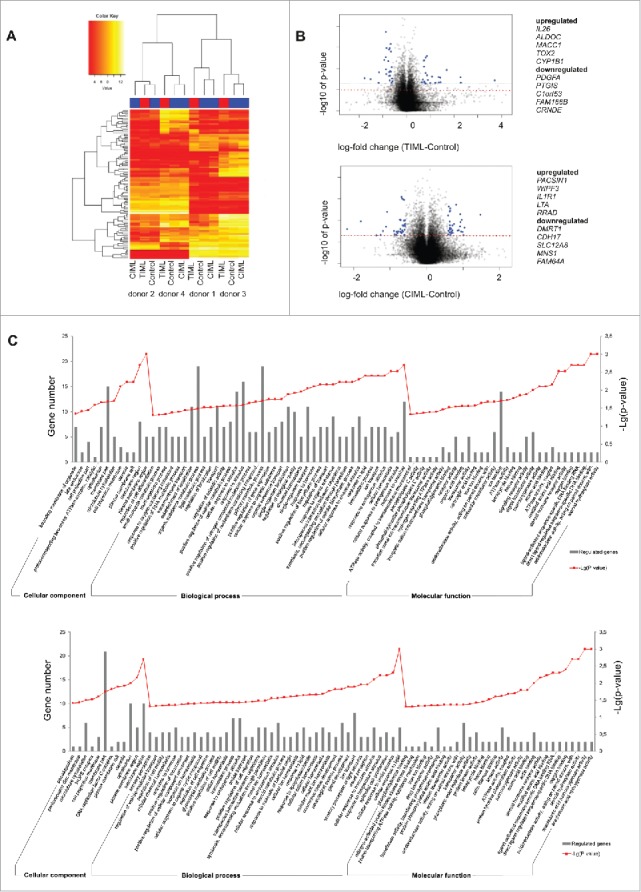
Fascinating earlier evidence suggests an intrinsic capacity of human natural killer (NK) cells to acquire adaptive immune features in the context of cytomegalovirus (CMV) infection or pro-inflammatory cytokine stimulation. Since the role of memory NK cells in cancer has so far remained elusive and adoptive NK cell transfer in relapsing pediatric acute B cell precursor leukemia (BCP-ALL) patients awaits improvement, we asked the question whether tumor-priming could promote the generation of memory NK cells with enhanced graft-vs.-leukemia (GvL) reactivity. Here, we provide substantial evidence that priming of naive human NK cells with pediatric acute B cell leukemia or acute myeloid leukemia specimens induces a functional conversion to tumor-induced memory-like (TIML)-NK cells displaying a heightened tumor-specific cytotoxicity and enhanced perforin synthesis. Cell cycles analyses reveal that tumor-priming sustainably alters the balance between NK cell activation and apoptosis in favor of survival. In addition, gene expression patterns differ between TIML- and cytokine-induced memory-like (CIML)-NK cells with the magnitude of regulated genes being distinctly higher in TIML-NK cells. As such, the tumor-induced conversion of NK cells triggers the emergence of a so far unacknowledged NK cell differentiation stage that might promote GvL effects in the context of adoptive cell transfer.
Keywords: Acute B cell precursor leukemia; adoptive cell transfer; children; natural killer cells.
Figures
References
-
- Pfeiffer M, Schumm M, Feuchtinger T, Dietz K, Handgretinger R, Lang P. Intensity of HLA class I expression and KIR-mismatch determine NK-cell mediated lysis of leukaemic blasts from children with acute lymphatic leukaemia. Br J Haematol 2007; 138(1):97-100; PMID:17555452; https://doi.org/10.1111/j.1365-2141.2007.06631.x - DOI - PubMed
-
- Pende D, Spaggiari GM, Marcenaro S, Martini S, Rivera P, Capobianco A, Falco M, Lanino E, Pierri I, Zambello R et al. . Analysis of the receptor-ligand interactions in the natural killer-mediated lysis of freshly isolated myeloid or lymphoblastic leukemias: Evidence for the involvement of the Poliovirus receptor (CD155) and Nectin-2 (CD112). Blood 2005; 105(5):2066-73; PMID:15536144; https://doi.org/10.1182/blood-2004-09-3548 - DOI - PubMed
-
- Pende D, Marcenaro S, Falco M, Martini S, Bernardo ME, Montagna D, Romeo E, Cognet C, Martinetti M, Maccario R et al. . Anti-leukemia activity of alloreactive NK cells in KIR ligand-mismatched haploidentical HSCT for pediatric patients: Evaluation of the functional role of activating KIR and redefinition of inhibitory KIR specificity. Blood 2009; 113(13):3119-29; PMID:18945967; https://doi.org/10.1182/blood-2008-06-164103 - DOI - PubMed
-
- Kübler A, Woiterski J, Witte K-E, Bühring H-J, Hartwig UF, Ebinger M, Oevermann L, Mezger M, Herr W, Lang P et al. . Both mature KIR+ and immature KIR− NK cells control pediatric acute B-cell precursor leukemia in NOD.Cg-Prkdcscid IL2rgtmWjl/Sz mice. Blood 2014; 124(26):3914-23; PMID:25359989; https://doi.org/10.1182/blood-2014-05-572743 - DOI - PubMed
-
- Handgretinger R, Lang P, Andre MC. Exploitation of Natural Killer (NK) cells for the treatment of acute leukemia. Blood 2016; 127(26):3341-9; PMID:27207791; https://doi.org/10.1182/blood-2015-12-629055 - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials