

Three-Dimensional Spatiotemporal Modeling of Colon Cancer Organoids Reveals that Multimodal Control of Stem Cell Self-Renewal is a Critical Determinant of Size and Shape in Early Stages of Tumor Growth
- PMID: 28681151
- PMCID: PMC5756149
- DOI: 10.1007/s11538-017-0294-1
Three-Dimensional Spatiotemporal Modeling of Colon Cancer Organoids Reveals that Multimodal Control of Stem Cell Self-Renewal is a Critical Determinant of Size and Shape in Early Stages of Tumor Growth
Abstract
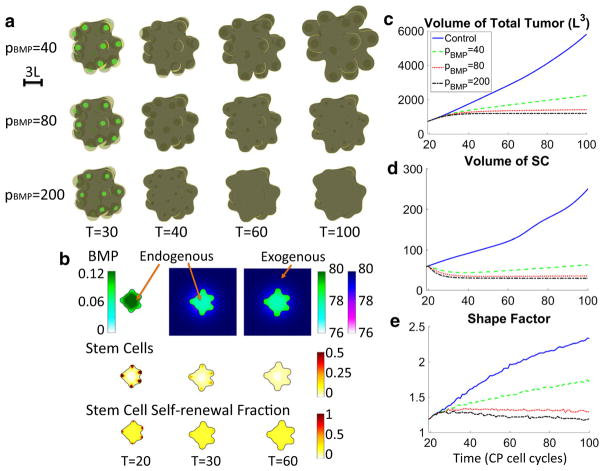
We develop a three-dimensional multispecies mathematical model to simulate the growth of colon cancer organoids containing stem, progenitor and terminally differentiated cells, as a model of early (prevascular) tumor growth. Stem cells (SCs) secrete short-range self-renewal promoters (e.g., Wnt) and their long-range inhibitors (e.g., Dkk) and proliferate slowly. Committed progenitor (CP) cells proliferate more rapidly and differentiate to produce post-mitotic terminally differentiated cells that release differentiation promoters, forming negative feedback loops on SC and CP self-renewal. We demonstrate that SCs play a central role in normal and cancer colon organoids. Spatial patterning of the SC self-renewal promoter gives rise to SC clusters, which mimic stem cell niches, around the organoid surface, and drive the development of invasive fingers. We also study the effects of externally applied signaling factors. Applying bone morphogenic proteins, which inhibit SC and CP self-renewal, reduces invasiveness and organoid size. Applying hepatocyte growth factor, which enhances SC self-renewal, produces larger sizes and enhances finger development at low concentrations but suppresses fingers at high concentrations. These results are consistent with recent experiments on colon organoids. Because many cancers are hierarchically organized and are subject to feedback regulation similar to that in normal tissues, our results suggest that in cancer, control of cancer stem cell self-renewal should influence the size and shape in similar ways, thereby opening the door to novel therapies.
Keywords: Brain tumors; Cancer stem cells; Cancer therapies; Feedback regulation; Mathematical modeling.
Figures











Similar articles
-
Multispecies model of cell lineages and feedback control in solid tumors.J Theor Biol. 2012 Jul 7;304:39-59. doi: 10.1016/j.jtbi.2012.02.030. Epub 2012 Mar 31. J Theor Biol. 2012. PMID: 22554945 Free PMC article.
-
FGF Signalling in the Self-Renewal of Colon Cancer Organoids.Sci Rep. 2019 Nov 22;9(1):17365. doi: 10.1038/s41598-019-53907-7. Sci Rep. 2019. PMID: 31758153 Free PMC article.
-
FGF2 and EGF Are Required for Self-Renewal and Organoid Formation of Canine Normal and Tumor Breast Stem Cells.J Cell Biochem. 2017 Mar;118(3):570-584. doi: 10.1002/jcb.25737. Epub 2016 Nov 28. J Cell Biochem. 2017. PMID: 27632571
-
Self-renewal molecular mechanisms of colorectal cancer stem cells.Int J Mol Med. 2017 Jan;39(1):9-20. doi: 10.3892/ijmm.2016.2815. Epub 2016 Nov 30. Int J Mol Med. 2017. PMID: 27909729 Free PMC article. Review.
-
Human colon cancer stem cells: a new paradigm in gastrointestinal oncology.J Clin Oncol. 2008 Jun 10;26(17):2828-38. doi: 10.1200/JCO.2008.17.6941. J Clin Oncol. 2008. PMID: 18539961 Review.
Cited by
-
Making human pancreatic islet organoids: Progresses on the cell origins, biomaterials and three-dimensional technologies.Theranostics. 2022 Jan 3;12(4):1537-1556. doi: 10.7150/thno.66670. eCollection 2022. Theranostics. 2022. PMID: 35198056 Free PMC article. Review.
-
Mathematical Models of Organoid Cultures.Front Genet. 2019 Sep 19;10:873. doi: 10.3389/fgene.2019.00873. eCollection 2019. Front Genet. 2019. PMID: 31592020 Free PMC article. Review.
-
Organoid Cultures In Silico: Tools or Toys?Bioengineering (Basel). 2022 Dec 30;10(1):50. doi: 10.3390/bioengineering10010050. Bioengineering (Basel). 2022. PMID: 36671623 Free PMC article. Review.
-
Cahn-Hilliard dynamical models for condensed biomolecular systems.bioRxiv [Preprint]. 2025 Jul 17:2025.07.13.664571. doi: 10.1101/2025.07.13.664571. bioRxiv. 2025. PMID: 40791366 Free PMC article. Preprint.
-
Simulations of tumor growth and response to immunotherapy by coupling a spatial agent-based model with a whole-patient quantitative systems pharmacology model.PLoS Comput Biol. 2022 Jul 22;18(7):e1010254. doi: 10.1371/journal.pcbi.1010254. eCollection 2022 Jul. PLoS Comput Biol. 2022. PMID: 35867773 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
