

Conservation and divergence of mitochondrial apoptosis pathway in the Pacific oyster, Crassostrea gigas
- PMID: 28682310
- PMCID: PMC5550854
- DOI: 10.1038/cddis.2017.307
Conservation and divergence of mitochondrial apoptosis pathway in the Pacific oyster, Crassostrea gigas
Abstract
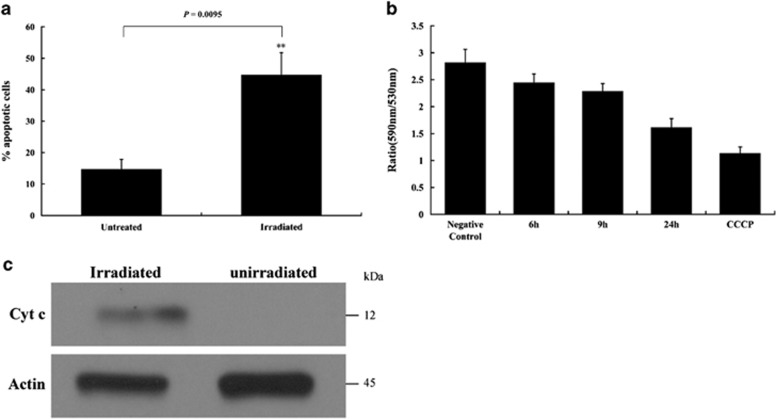
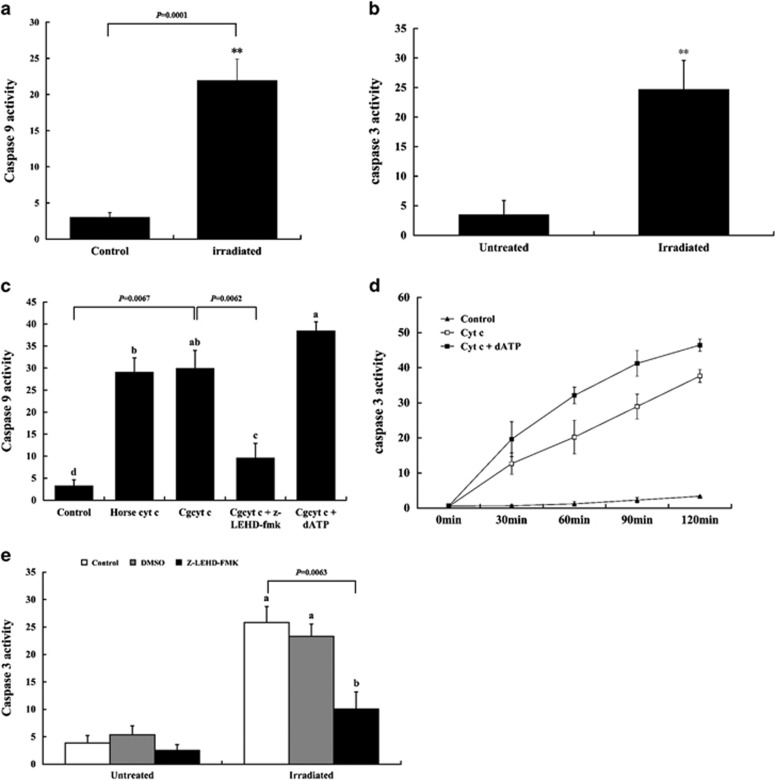
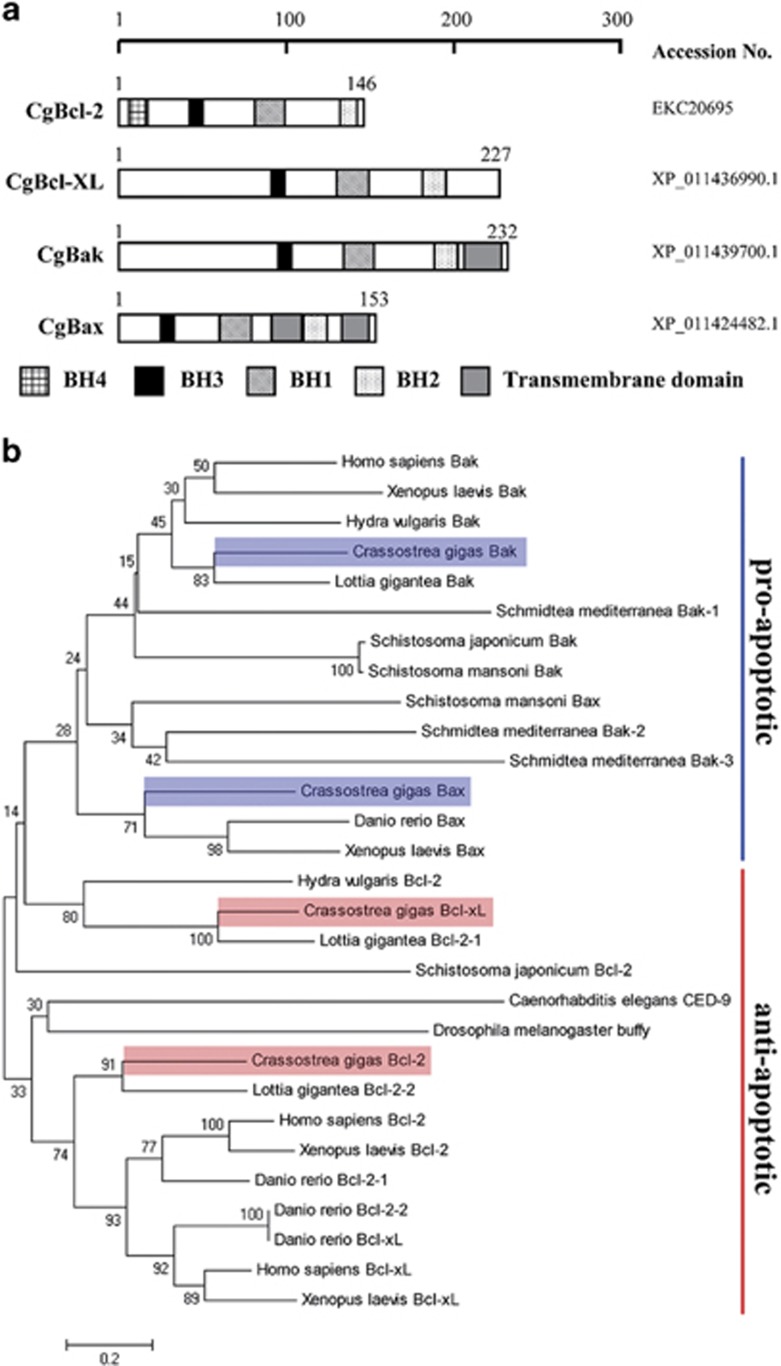
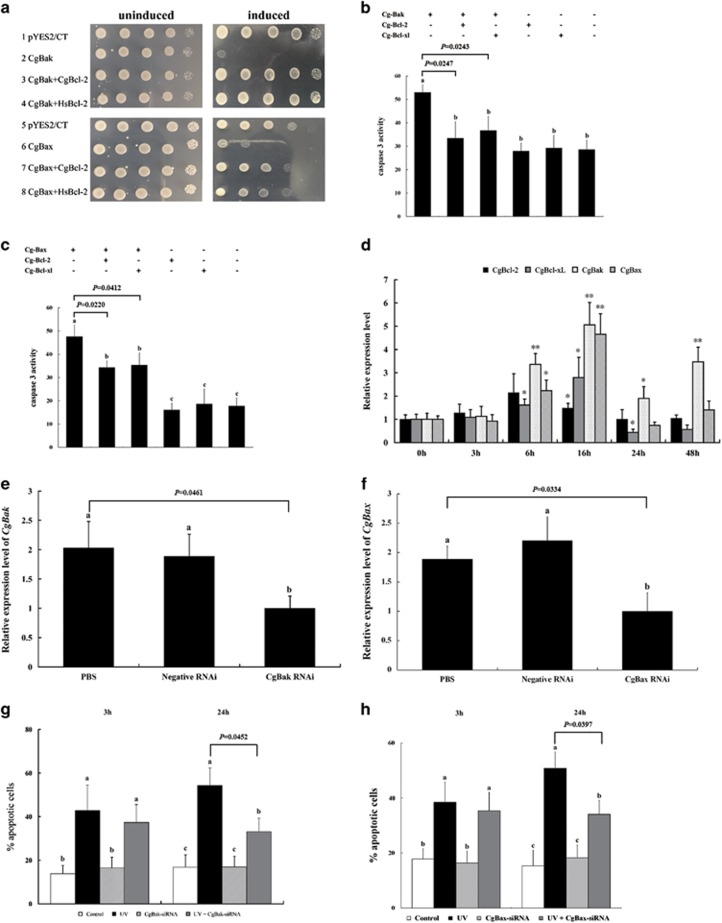
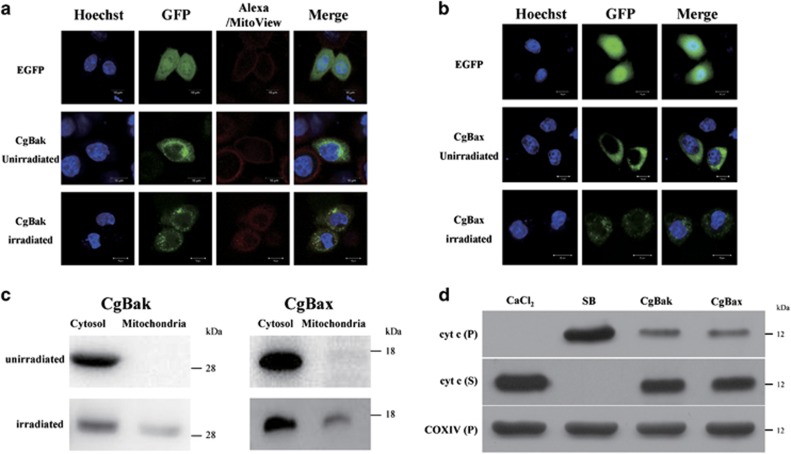
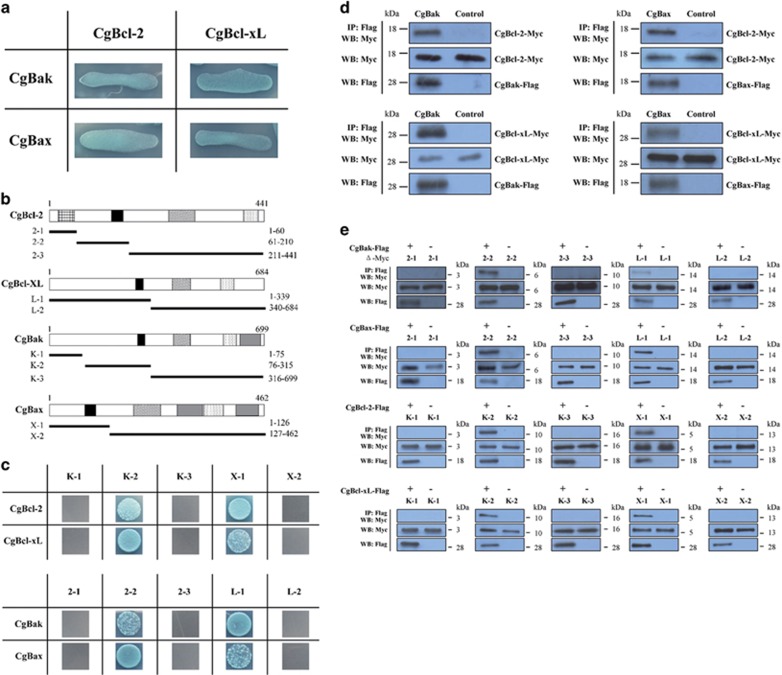
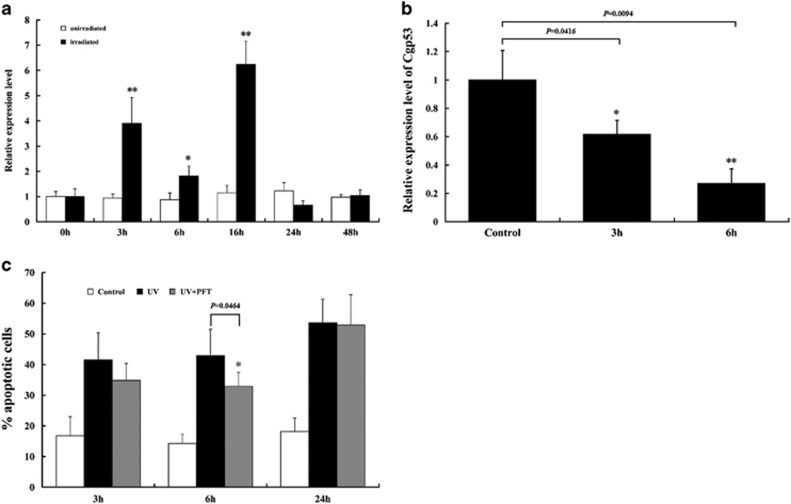
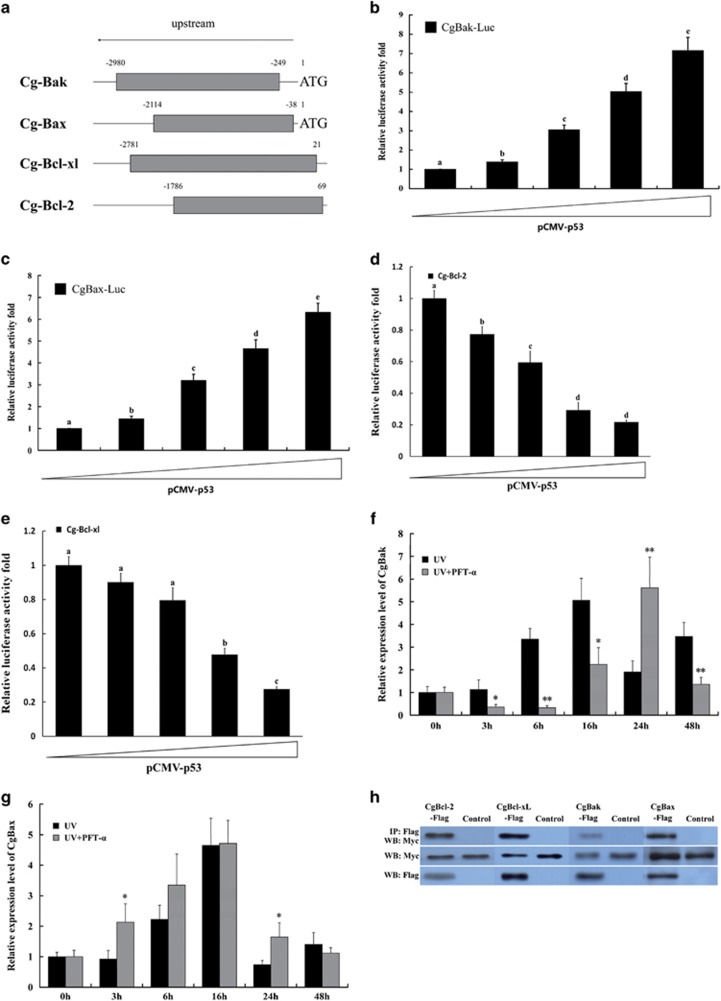
Apoptosis is considered a crucial part of the host defense system in oysters according to previous reports; however, the exact process by which this occurs remains unclear. Besides, mitochondrial apoptosis is the primary method of apoptosis in vertebrate cells, but has been poorly studied in invertebrates and is quite controversial. In this study, we investigated the molecular mechanism of mitochondrial apoptosis in the Pacific oyster Crassostrea gigas. Notably, we show that most key elements involved in the vertebrate mitochondrial apoptosis pathway - including mitochondrial outer membrane permeabilization, cytochrome c release, and caspase activation - are also present in C. gigas. In contrast, the lack of Bcl-2 homology 3-only subfamily members and apoptotic protease activating factor-1 (APAF-1) protein revealed evolutionary diversity from other phyla. Our results support that mitochondrial apoptosis in animals predates the emergence of vertebrates, but suggest that an unexpectedly diverse mitochondrial apoptosis pathway may exist in invertebrates. In addition, our work provided new clues for an improved understanding of how bivalve acclimate themselves to an inconstant environment.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Changes in intramitochondrial and cytosolic pH: early events that modulate caspase activation during apoptosis.Nat Cell Biol. 2000 Jun;2(6):318-25. doi: 10.1038/35014006. Nat Cell Biol. 2000. PMID: 10854321
-
The modulation of Smac/DIABLO on mitochondrial apoptosis induced by LPS in Crassostrea gigas.Fish Shellfish Immunol. 2019 Jan;84:587-598. doi: 10.1016/j.fsi.2018.10.035. Epub 2018 Oct 15. Fish Shellfish Immunol. 2019. PMID: 30336283
-
Connected to death: the (unexpurgated) mitochondrial pathway of apoptosis.Science. 2005 Oct 7;310(5745):66-7. doi: 10.1126/science.1117105. Science. 2005. PMID: 16210526
-
Multiple pathways of cytochrome c release from mitochondria in apoptosis.Biochim Biophys Acta. 2006 May-Jun;1757(5-6):639-47. doi: 10.1016/j.bbabio.2006.03.016. Epub 2006 Apr 19. Biochim Biophys Acta. 2006. PMID: 16678785 Review.
-
Mechanisms of cytochrome c release from mitochondria.Cell Death Differ. 2006 Sep;13(9):1423-33. doi: 10.1038/sj.cdd.4401950. Epub 2006 May 5. Cell Death Differ. 2006. PMID: 16676004 Review.
Cited by
-
Characterization of the far-red fluorescent probe MitoView 633 for dynamic mitochondrial membrane potential measurement.Front Physiol. 2023 Oct 23;14:1257739. doi: 10.3389/fphys.2023.1257739. eCollection 2023. Front Physiol. 2023. PMID: 37936577 Free PMC article.
-
GABA mitigates mitochondrial apoptosis induced by high temperature stress in the Pacific oyster (Crassostrea gigas).Mol Biol Rep. 2025 Jan 22;52(1):147. doi: 10.1007/s11033-025-10238-3. Mol Biol Rep. 2025. PMID: 39841302
-
A novel Fas ligand plays an important role in cell apoptosis of Crassostrea hongkongensis: molecular cloning, expression profiles and functional identification of ChFasL.Front Immunol. 2023 Oct 5;14:1267772. doi: 10.3389/fimmu.2023.1267772. eCollection 2023. Front Immunol. 2023. PMID: 37868973 Free PMC article.
-
Status in molluscan cell line development in last one decade (2010-2020): impediments and way forward.Cytotechnology. 2022 Aug;74(4):433-457. doi: 10.1007/s10616-022-00539-x. Epub 2022 Jul 24. Cytotechnology. 2022. PMID: 36110153 Free PMC article. Review.
-
Phylogenetic analysis of the caspase family in bivalves: implications for programmed cell death, immune response and development.BMC Genomics. 2021 Jan 25;22(1):80. doi: 10.1186/s12864-021-07380-0. BMC Genomics. 2021. PMID: 33494703 Free PMC article.
References
-
- Guofan Z, Xiaodong F, Ximing G, Li L, Ruibang L, Fei X et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012; 490: 49–54. - PubMed
-
- Francis MH, Brent F, Snimar G, Inna MS. Apoptosis as a host defense mechanism in Crassostrea virginica and its modulation by Perkinsus marinus. Fish Shellfish Immunol 2010; 29: 247–257. - PubMed
-
- Sokolova IM, Evans S, Hughes FM. Cadmium-induced apoptosis in oyster hemocytes involves disturbance of cellular energy balance but no mitochondrial permeability transition. J Exp Biol 2004; 207: 3369–3380. - PubMed
-
- Foster B, Grewal S, Graves O, Hughes FM, Sokolova IM. Copper exposure affects hemocyte apoptosis and Perkinsus marinus infection in eastern oysters Crassostrea virginica (Gmelin). Fish Shellfish Immunol 2011; 31: 341–349. - PubMed
-
- Renault T, Faury N, Barbosa-Solomieu V, Moreau K. Suppression substractive hybridisation (SSH) and real time PCR reveal differential gene expression in the Pacific cupped oyster, Crassostrea gigas, challenged with Ostreid herpesvirus 1. Dev Comp Immunol 2011; 35: 725–735. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
