

Toward a Neurocentric View of Learning
- PMID: 28683265
- PMCID: PMC5519140
- DOI: 10.1016/j.neuron.2017.05.021
Toward a Neurocentric View of Learning
Abstract
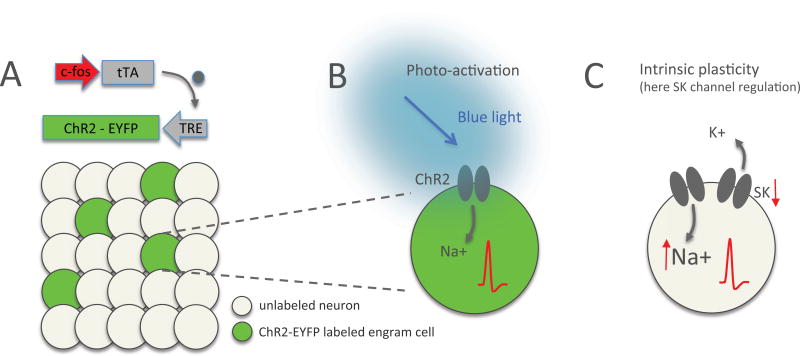
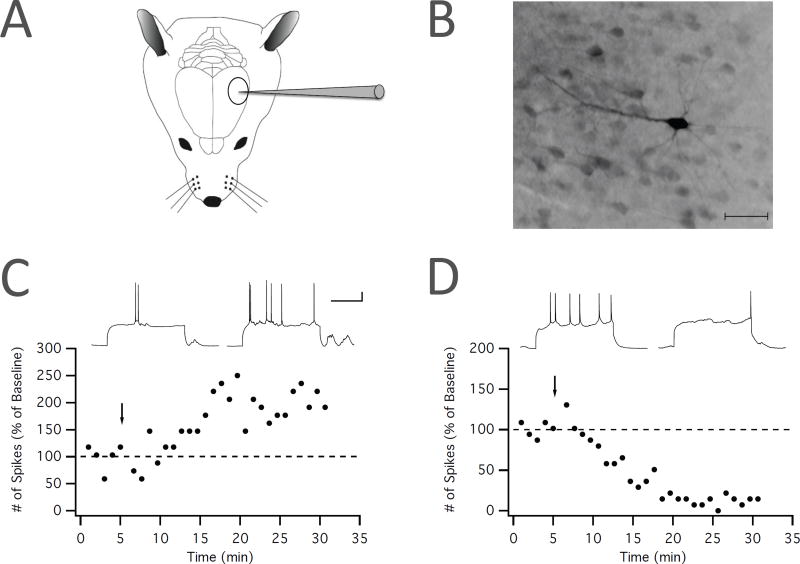
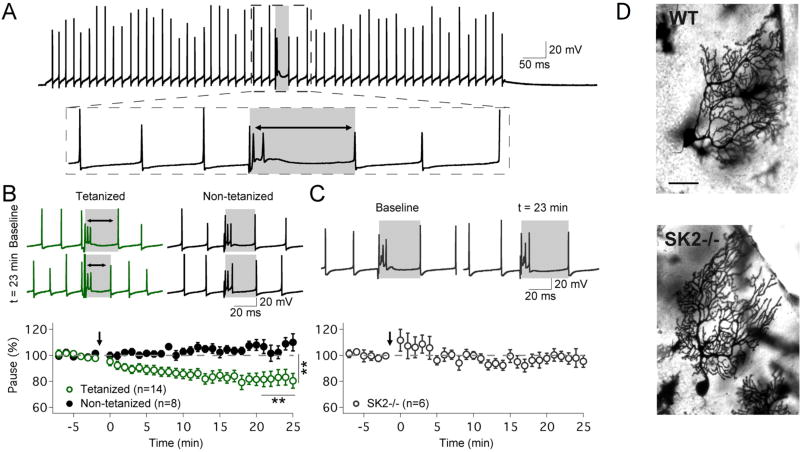
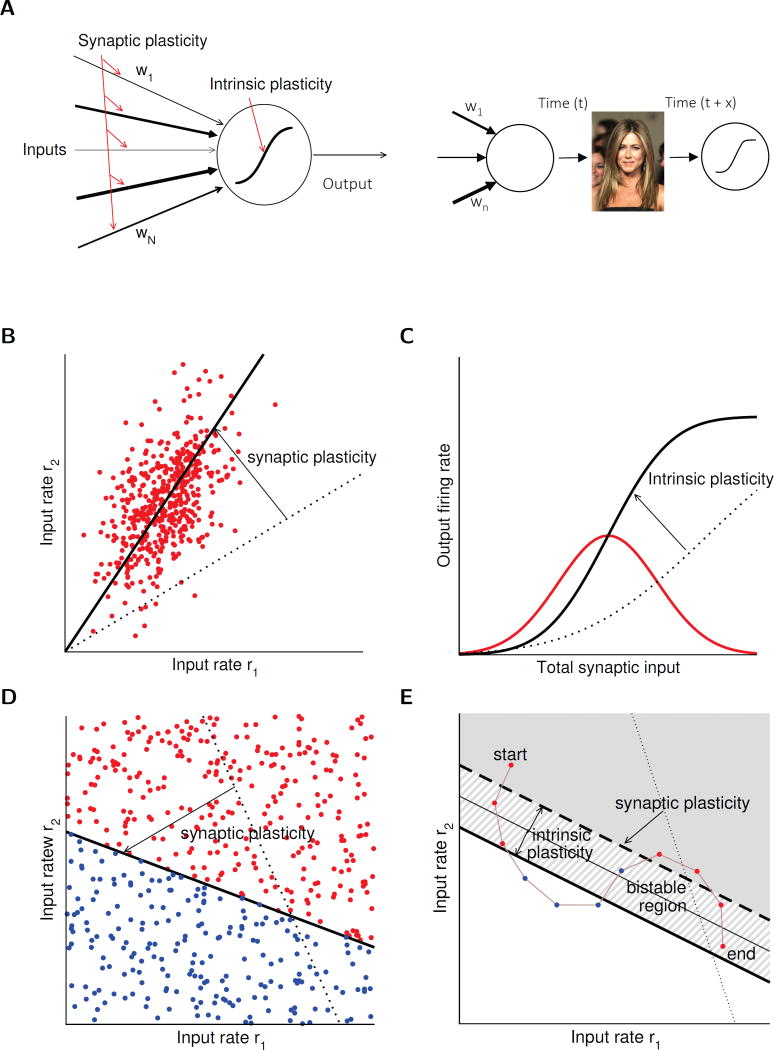
Synaptic plasticity (e.g., long-term potentiation [LTP]) is considered the cellular correlate of learning. Recent optogenetic studies on memory engram formation assign a critical role in learning to suprathreshold activation of neurons and their integration into active engrams ("engram cells"). Here we review evidence that ensemble integration may result from LTP but also from cell-autonomous changes in membrane excitability. We propose that synaptic plasticity determines synaptic connectivity maps, whereas intrinsic plasticity-possibly separated in time-amplifies neuronal responsiveness and acutely drives engram integration. Our proposal marks a move away from an exclusively synaptocentric toward a non-exclusive, neurocentric view of learning.
Keywords: Purkinje cell; cerebellum; ensemble; hippocampus; intrinsic; memory engram; neocortex; plasticity; pyramidal cell; synaptic.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Synergistic excitability plasticity in cerebellar functioning.FEBS J. 2020 Nov;287(21):4557-4593. doi: 10.1111/febs.15355. Epub 2020 Jun 11. FEBS J. 2020. PMID: 32367676 Review.
-
Long-term plasticity of intrinsic excitability: learning rules and mechanisms.Learn Mem. 2003 Nov-Dec;10(6):456-65. doi: 10.1101/lm.64103. Learn Mem. 2003. PMID: 14657257 Review.
-
Properties and mechanisms of long-term synaptic plasticity in the mammalian brain: relationships to learning and memory.Neurobiol Learn Mem. 1995 Jan;63(1):1-18. doi: 10.1006/nlme.1995.1001. Neurobiol Learn Mem. 1995. PMID: 7663875 Review.
-
Learning-induced bidirectional enhancement of inhibitory synaptic metaplasticity.J Physiol. 2024 May;602(10):2343-2358. doi: 10.1113/JP284761. Epub 2024 Apr 23. J Physiol. 2024. PMID: 38654583
-
[The general principles of synaptic plasticity in the neocortex, hippocampus and cerebellum].Zh Vyssh Nerv Deiat Im I P Pavlova. 1997 Mar-Apr;47(2):374-92. Zh Vyssh Nerv Deiat Im I P Pavlova. 1997. PMID: 9173742 Review. Russian.
Cited by
-
Intracerebellar cannabinoid administration impairs delay but not trace eyeblink conditioning.Behav Brain Res. 2020 Jan 27;378:112258. doi: 10.1016/j.bbr.2019.112258. Epub 2019 Sep 24. Behav Brain Res. 2020. PMID: 31560921 Free PMC article.
-
Stable memory with unstable synapses.Nat Commun. 2019 Sep 30;10(1):4441. doi: 10.1038/s41467-019-12306-2. Nat Commun. 2019. PMID: 31570719 Free PMC article.
-
Transcranial direct current stimulation of cerebellum alters spiking precision in cerebellar cortex: A modeling study of cellular responses.PLoS Comput Biol. 2021 Dec 9;17(12):e1009609. doi: 10.1371/journal.pcbi.1009609. eCollection 2021 Dec. PLoS Comput Biol. 2021. PMID: 34882680 Free PMC article.
-
Reorganization of Recurrent Layer 5 Corticospinal Networks Following Adult Motor Training.J Neurosci. 2019 Jun 12;39(24):4684-4693. doi: 10.1523/JNEUROSCI.3442-17.2019. Epub 2019 Apr 4. J Neurosci. 2019. PMID: 30948479 Free PMC article.
-
Defective memory engram reactivation underlies impaired fear memory recall in Fragile X syndrome.Elife. 2020 Nov 20;9:e61882. doi: 10.7554/eLife.61882. Elife. 2020. PMID: 33215988 Free PMC article.
References
-
- Aizenman CD, Linden DJ. Rapid, synaptically driven increases in the intrinsic excitability of cerebellar deep nuclear neurons. Nat. Neurosci. 2000;3:109–111. - PubMed
-
- Albus JS. A theory of cerebellar function. Math. Biosciences. 1971;10:25–61.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
