

Ancillary Activity: Beyond Core Metabolism in Immune Cells
- PMID: 28683280
- PMCID: PMC5546226
- DOI: 10.1016/j.cmet.2017.06.019
Ancillary Activity: Beyond Core Metabolism in Immune Cells
Abstract
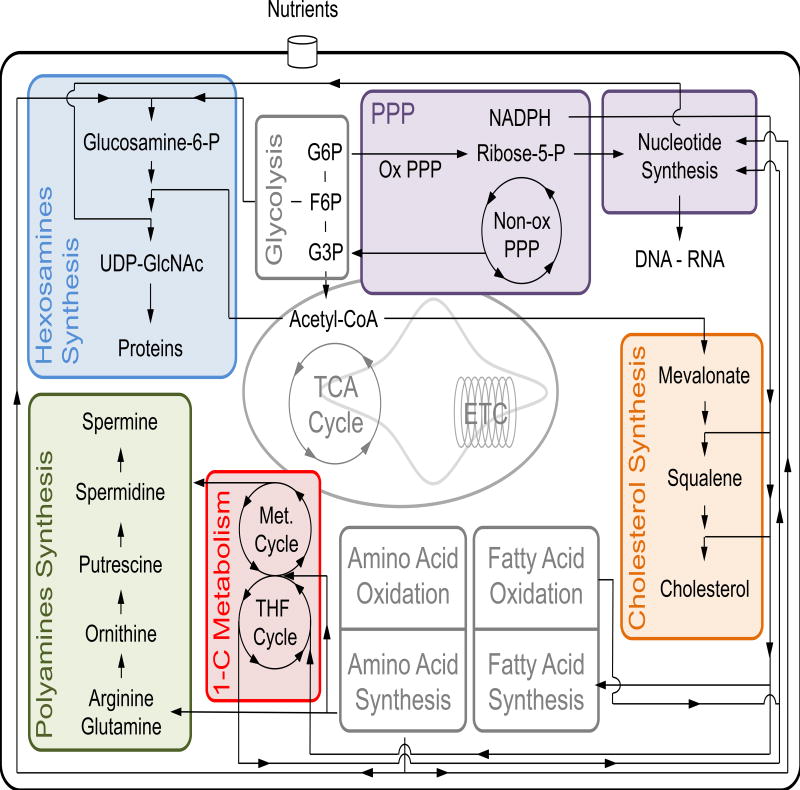
Immune cell function and fate are intimately linked to engagement of metabolic pathways. The contribution of core metabolic pathways to immune cell bioenergetics has been vigorously investigated in recent years. However, precisely how other peripheral metabolic pathways support immune cells beyond energy generation is less well understood. Here we survey the literature and highlight recent advances in our understanding of several ancillary metabolic pathways and how they support processes beyond ATP production and ultimately contribute to protective immunity.
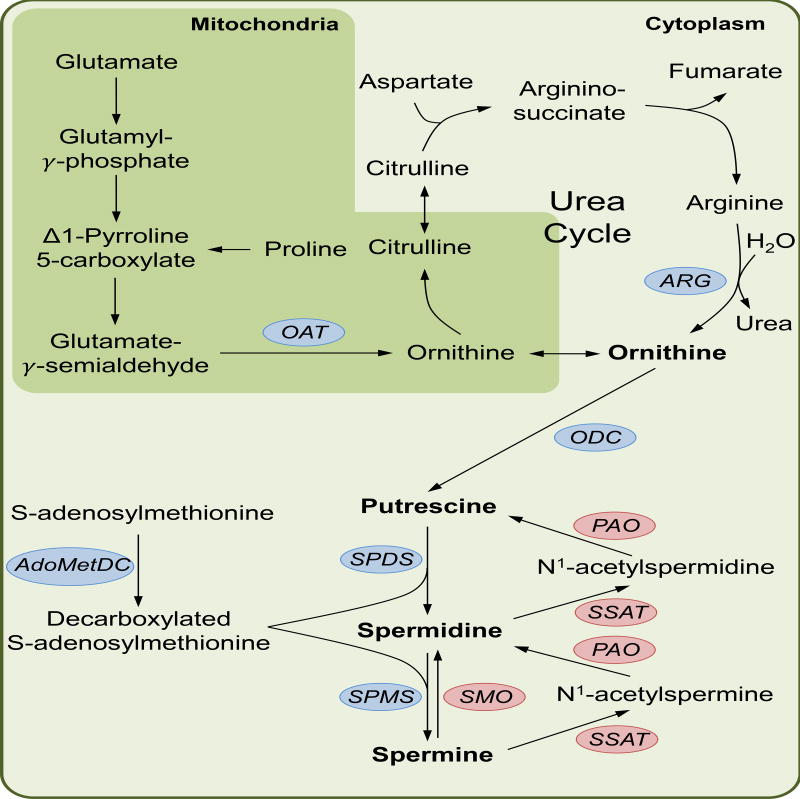
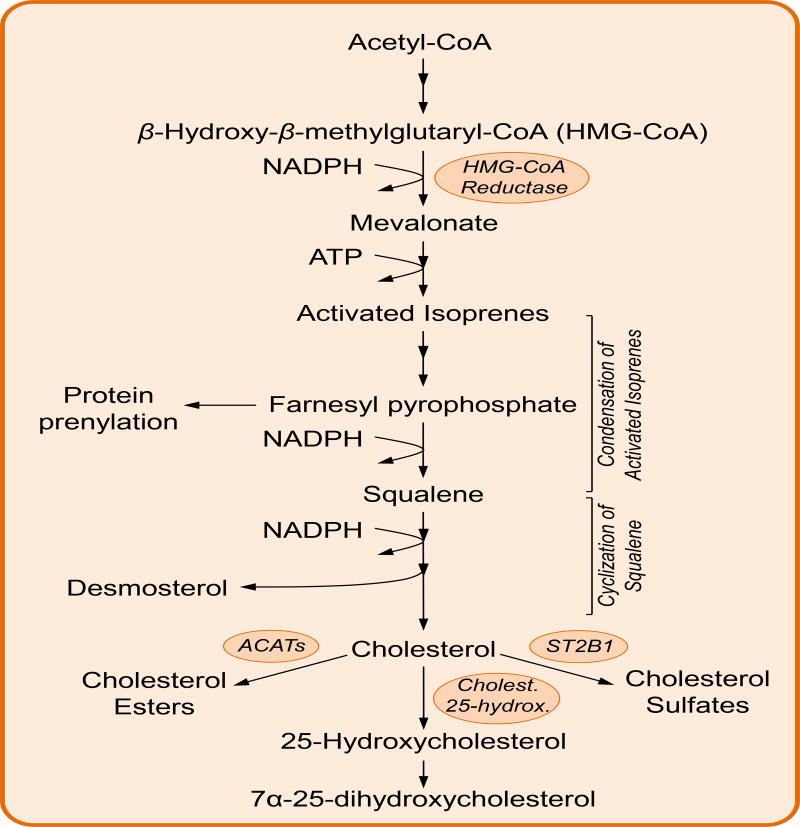
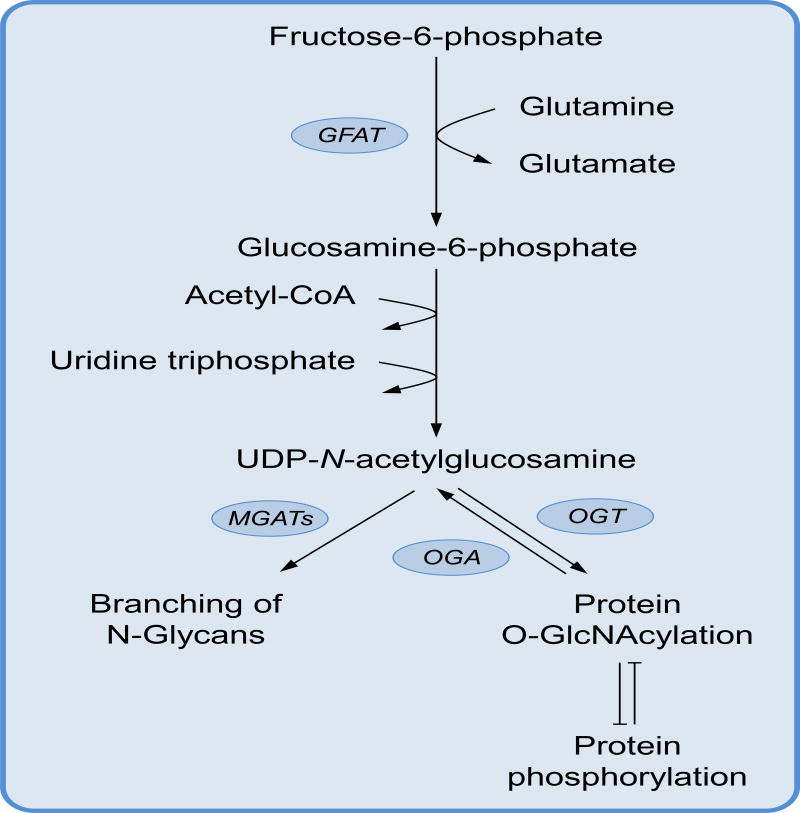
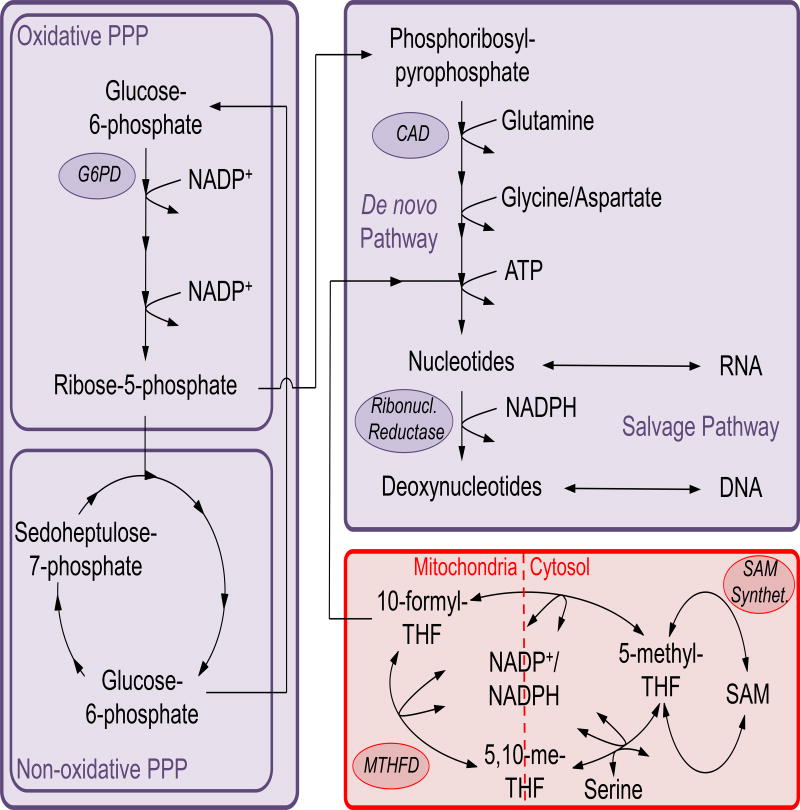
Keywords: O-GlcNAcylation; cholesterol; hexosamine synthesis pathway; immune cell metabolism; immunity; metabolism; nucleotides; one-carbon metabolism; pentose phosphate pathway; polyamines.
Copyright © 2017. Published by Elsevier Inc.
Figures
References
-
- Abdulhussein AA, Wallace HM. Polyamines and membrane transporters. Amino Acids. 2014;46:655–660. - PubMed
-
- Austin WR, Armijo AL, Campbell DO, Singh AS, Hsieh T, Nathanson D, Herschman HR, Phelps ME, Witte ON, Czernin J, et al. Nucleoside salvage pathway kinases regulate hematopoiesis by linking nucleotide metabolism with replication stress. Journal of Experimental Medicine. 2012;209:2215–2228. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
