

Silent Allosteric Modulation of mGluR5 Maintains Glutamate Signaling while Rescuing Alzheimer's Mouse Phenotypes
- PMID: 28683325
- PMCID: PMC5547898
- DOI: 10.1016/j.celrep.2017.06.023
Silent Allosteric Modulation of mGluR5 Maintains Glutamate Signaling while Rescuing Alzheimer's Mouse Phenotypes
Abstract
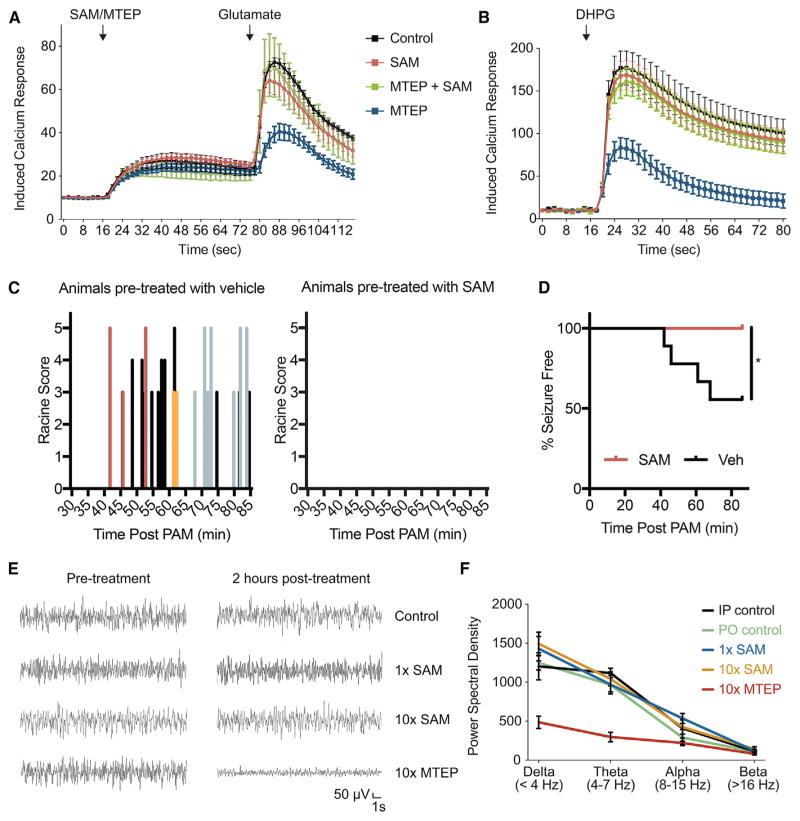
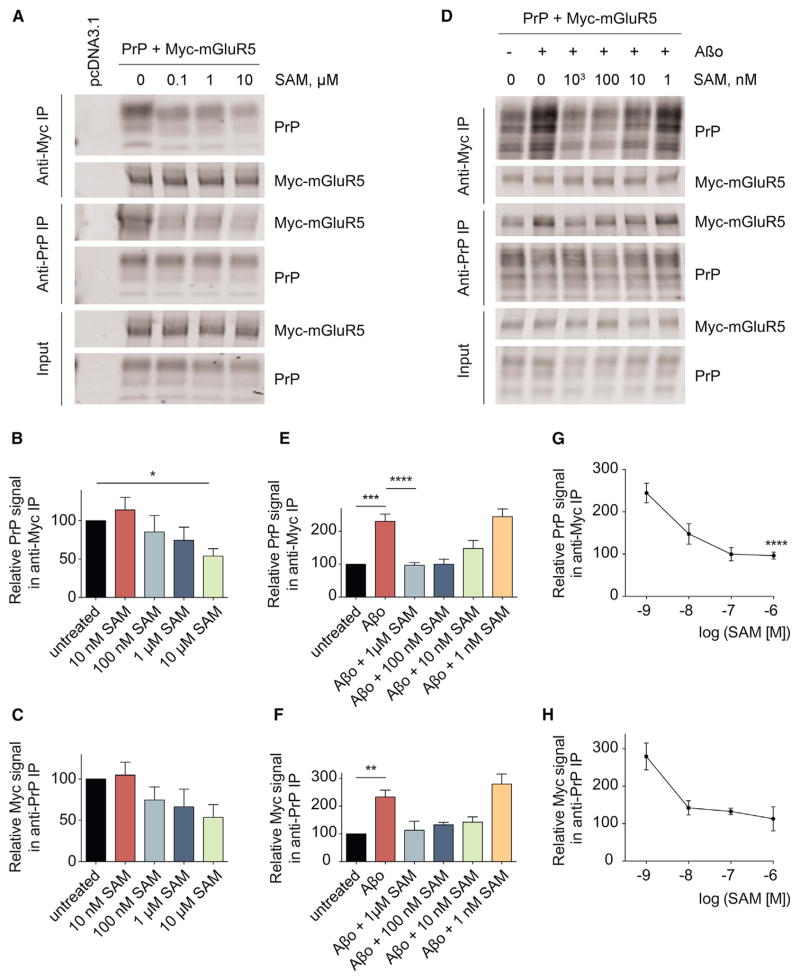
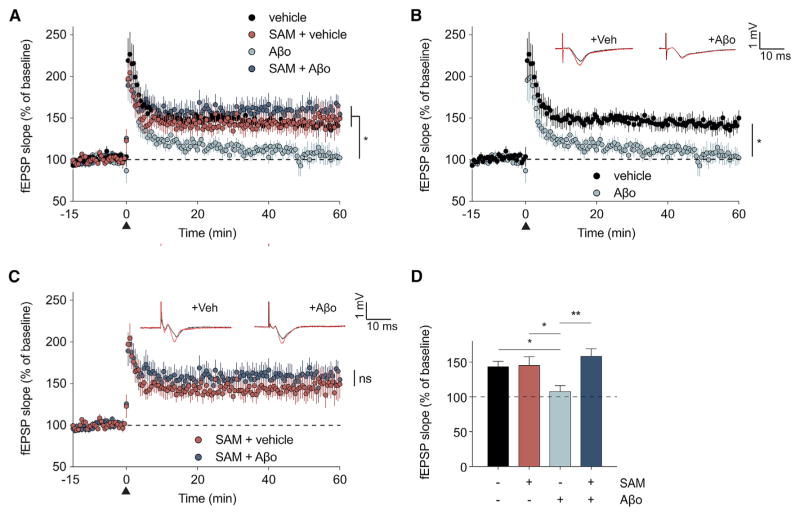
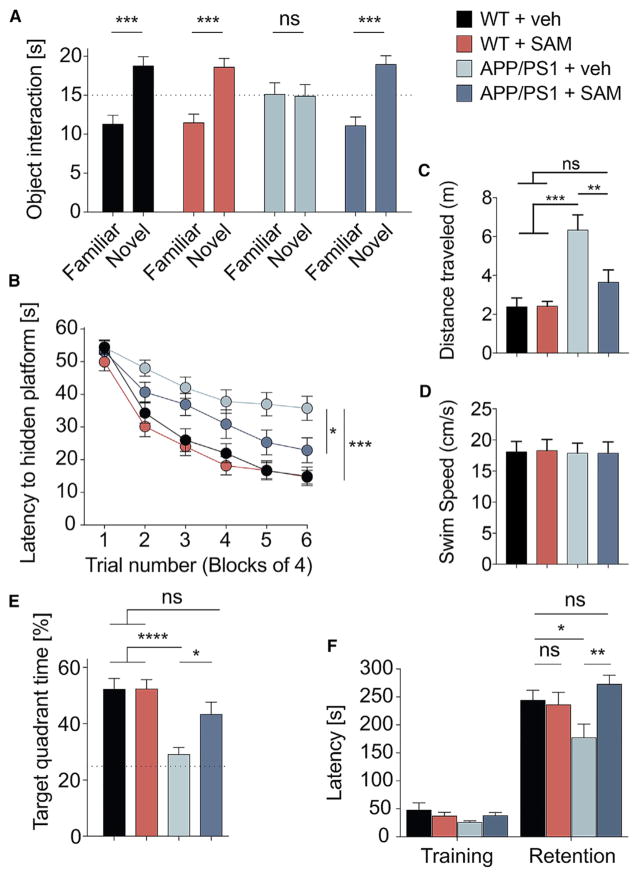
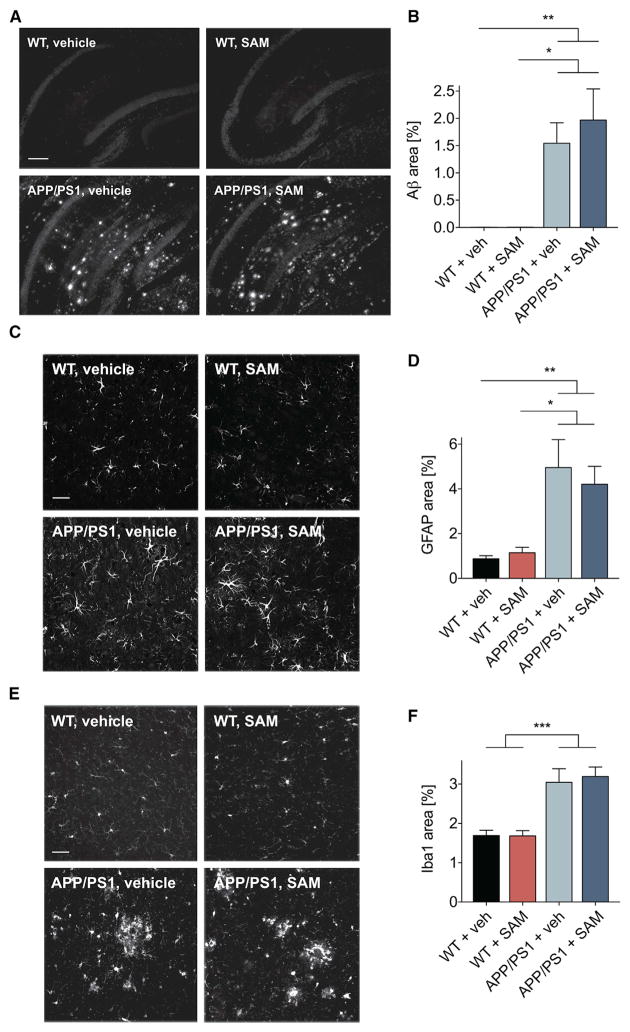
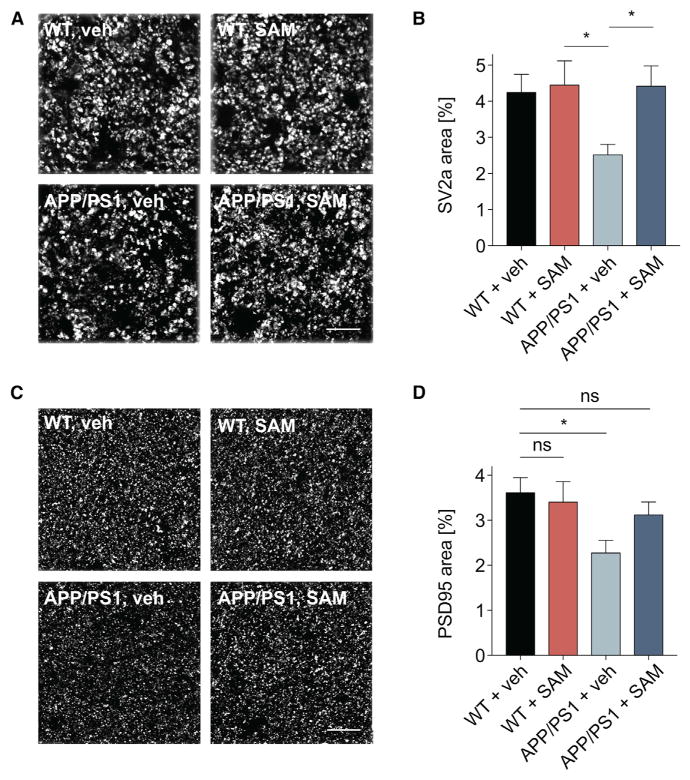
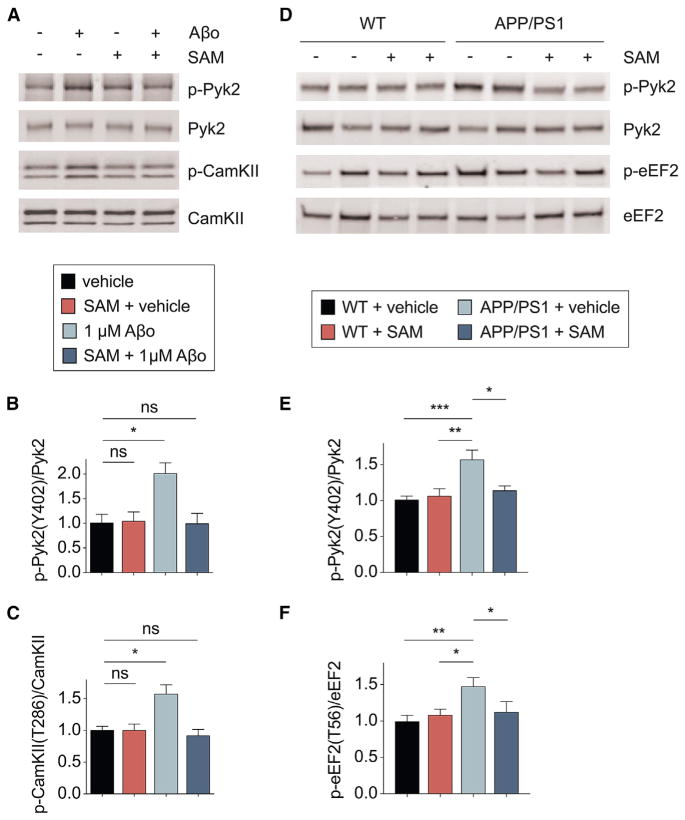
Metabotropic glutamate receptor 5 (mGluR5) has been implicated in Alzheimer's disease (AD) pathology. We sought to understand whether mGluR5's role in AD requires glutamate signaling. We used a potent mGluR5 silent allosteric modulator (SAM, BMS-984923) to separate its well-known physiological role in glutamate signaling from a pathological role in mediating amyloid-β oligomer (Aβo) action. Binding of the SAM to mGluR5 does not change glutamate signaling but strongly reduces mGluR5 interaction with cellular prion protein (PrPC) bound to Aβo. The SAM compound prevents Aβo-induced signal transduction in brain slices and in an AD transgenic mouse model, the APPswe/PS1ΔE9 strain. Critically, 4 weeks of SAM treatment rescues memory deficits and synaptic depletion in the APPswe/PS1ΔE9 transgenic mouse brain. Our data show that mGluR5's role in Aβo-dependent AD phenotypes is separate from its role in glutamate signaling and silent allosteric modulation of mGluR5 has promise as a disease-modifying AD intervention with a broad therapeutic window.
Keywords: Alzheimer; amyloid β; mGluR5; metabotropic glutamate receptor 5; oligomer; prion protein; silent allosteric modulator; transgenic mouse.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Therapeutic molecules and endogenous ligands regulate the interaction between brain cellular prion protein (PrPC) and metabotropic glutamate receptor 5 (mGluR5).J Biol Chem. 2014 Oct 10;289(41):28460-77. doi: 10.1074/jbc.M114.584342. Epub 2014 Aug 22. J Biol Chem. 2014. PMID: 25148681 Free PMC article.
-
Oligomers of Amyloid β Prevent Physiological Activation of the Cellular Prion Protein-Metabotropic Glutamate Receptor 5 Complex by Glutamate in Alzheimer Disease.J Biol Chem. 2016 Aug 12;291(33):17112-21. doi: 10.1074/jbc.M116.720664. Epub 2016 Jun 20. J Biol Chem. 2016. PMID: 27325698 Free PMC article.
-
Metabotropic glutamate receptor 5 couples cellular prion protein to intracellular signalling in Alzheimer's disease.Brain. 2016 Feb;139(Pt 2):526-46. doi: 10.1093/brain/awv356. Epub 2015 Dec 14. Brain. 2016. PMID: 26667279 Free PMC article.
-
Noncanonical Metabotropic Glutamate Receptor 5 Signaling in Alzheimer's Disease.Annu Rev Pharmacol Toxicol. 2022 Jan 6;62:235-254. doi: 10.1146/annurev-pharmtox-021821-091747. Epub 2021 Sep 13. Annu Rev Pharmacol Toxicol. 2022. PMID: 34516293 Review.
-
Synaptotoxic Signaling by Amyloid Beta Oligomers in Alzheimer's Disease Through Prion Protein and mGluR5.Adv Pharmacol. 2018;82:293-323. doi: 10.1016/bs.apha.2017.09.007. Epub 2017 Oct 25. Adv Pharmacol. 2018. PMID: 29413525 Free PMC article. Review.
Cited by
-
A Role for Cellular Prion Protein in Late-Onset Alzheimer's Disease: Evidence from Preclinical Studies.J Neurosci. 2018 Feb 28;38(9):2146-2148. doi: 10.1523/JNEUROSCI.3307-17.2018. J Neurosci. 2018. PMID: 29491138 Free PMC article. Review. No abstract available.
-
Homer1a and mGluR1/5 Signaling in Homeostatic Sleep Drive and Output.Yale J Biol Med. 2019 Mar 25;92(1):93-101. eCollection 2019 Mar. Yale J Biol Med. 2019. PMID: 30923476 Free PMC article. Review.
-
An Expanded Narrative Review of Neurotransmitters on Alzheimer's Disease: The Role of Therapeutic Interventions on Neurotransmission.Mol Neurobiol. 2025 Feb;62(2):1631-1674. doi: 10.1007/s12035-024-04333-y. Epub 2024 Jul 16. Mol Neurobiol. 2025. PMID: 39012443 Free PMC article. Review.
-
Targeting synapse function and loss for treatment of neurodegenerative diseases.Nat Rev Drug Discov. 2024 Jan;23(1):23-42. doi: 10.1038/s41573-023-00823-1. Epub 2023 Nov 27. Nat Rev Drug Discov. 2024. PMID: 38012296 Review.
-
Molecular Pathogenesis and Interventional Strategies for Alzheimer's Disease: Promises and Pitfalls.ACS Pharmacol Transl Sci. 2020 Mar 26;3(3):472-488. doi: 10.1021/acsptsci.9b00104. eCollection 2020 Jun 12. ACS Pharmacol Transl Sci. 2020. PMID: 32566913 Free PMC article. Review.
References
-
- Ballatore C, Lee VM, Trojanowski JQ. Tau-mediated neurodegeneration in Alzheimer’s disease and related disorders. Nat Rev Neurosci. 2007;8:663–672. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
