

A Plastid Phosphatidylglycerol Lipase Contributes to the Export of Acyl Groups from Plastids for Seed Oil Biosynthesis
- PMID: 28687655
- PMCID: PMC5559756
- DOI: 10.1105/tpc.17.00397
A Plastid Phosphatidylglycerol Lipase Contributes to the Export of Acyl Groups from Plastids for Seed Oil Biosynthesis
Abstract
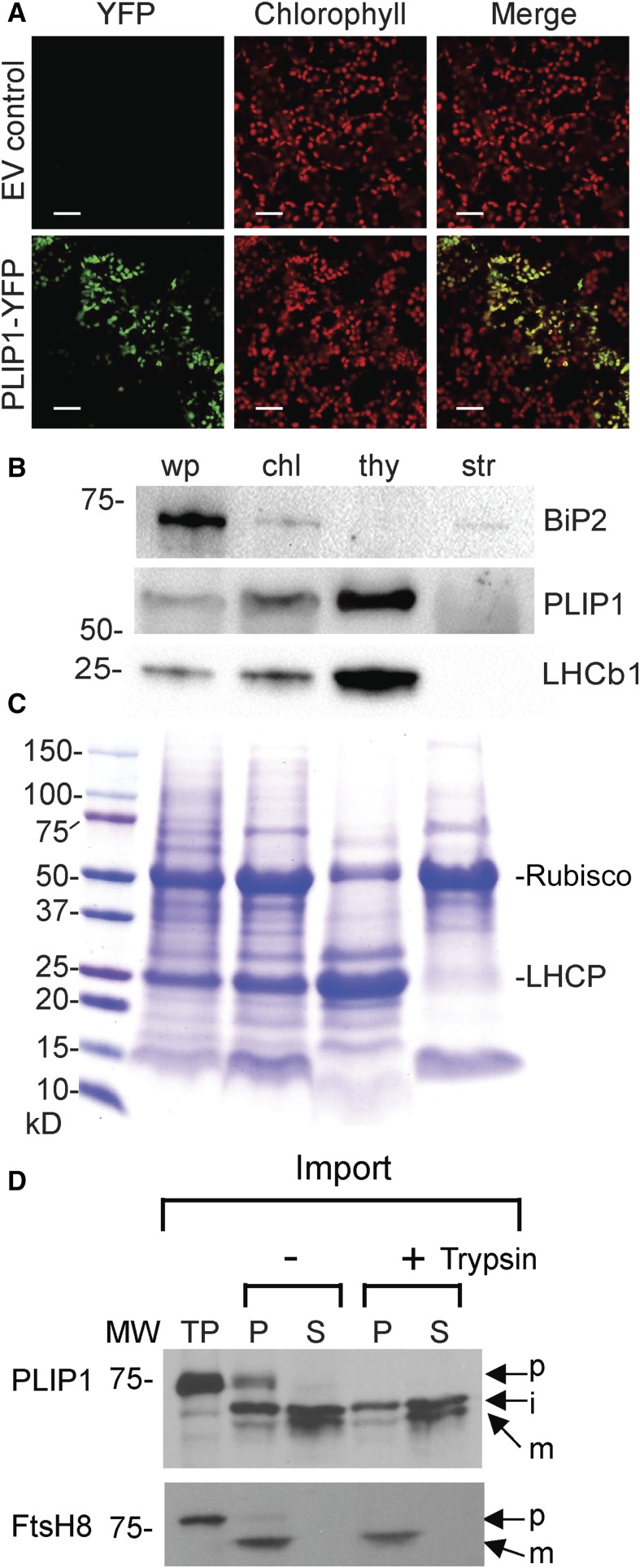
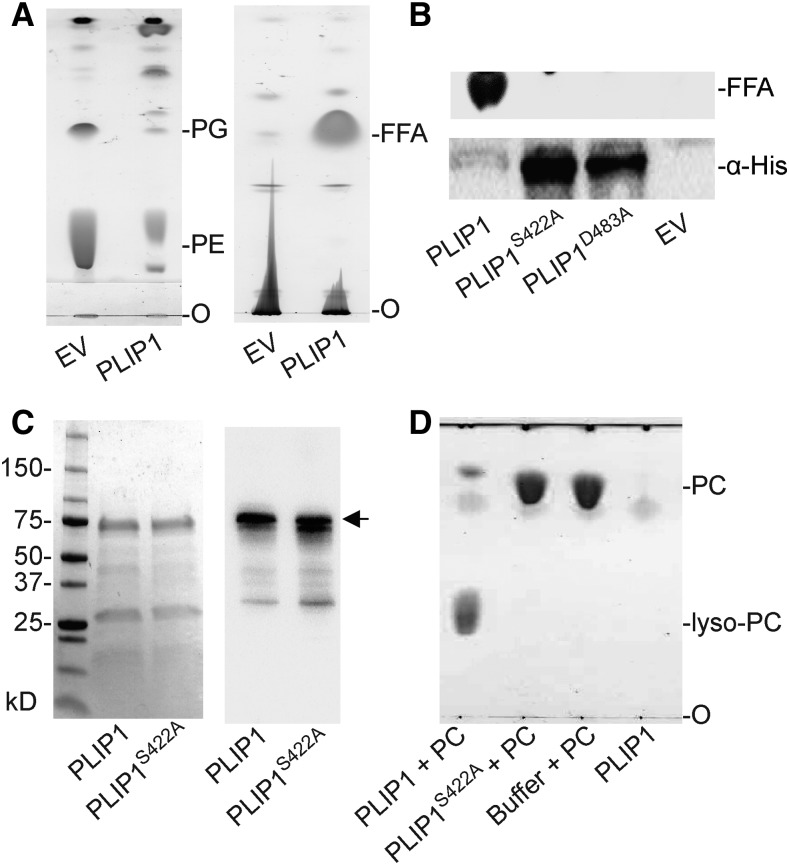
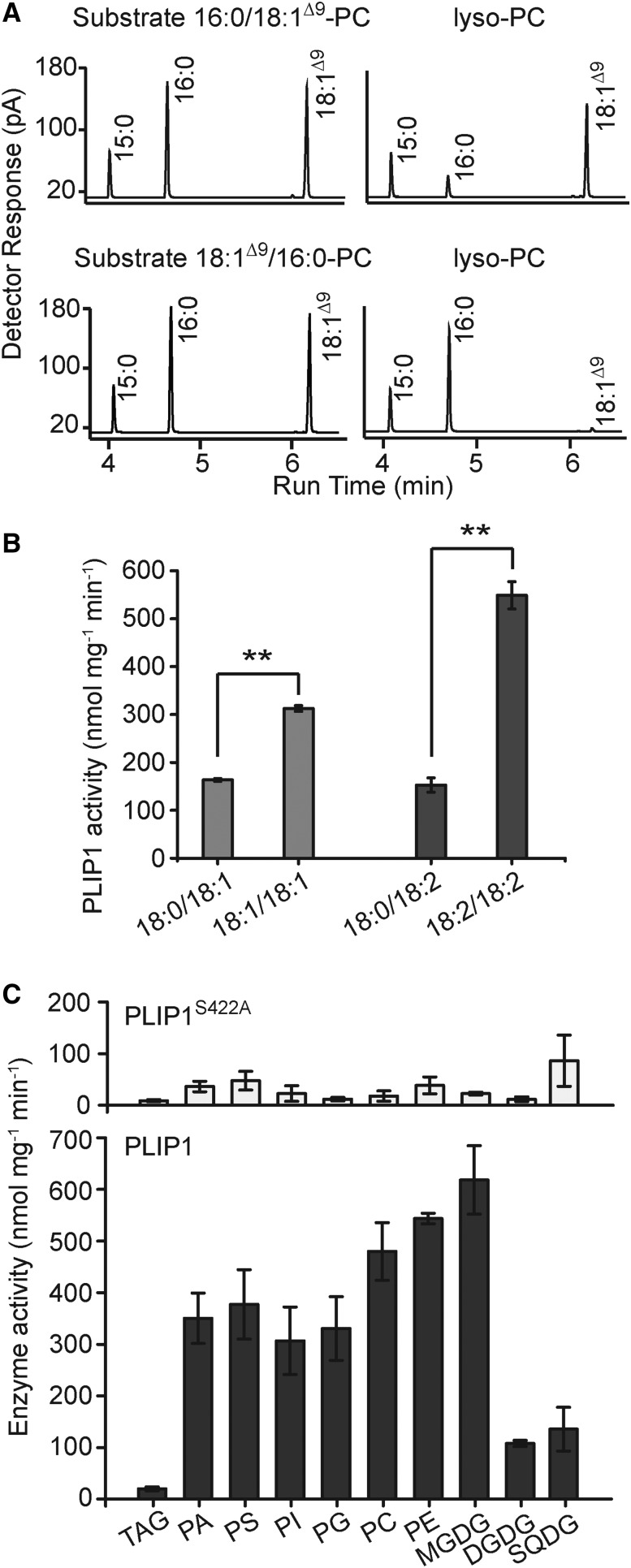
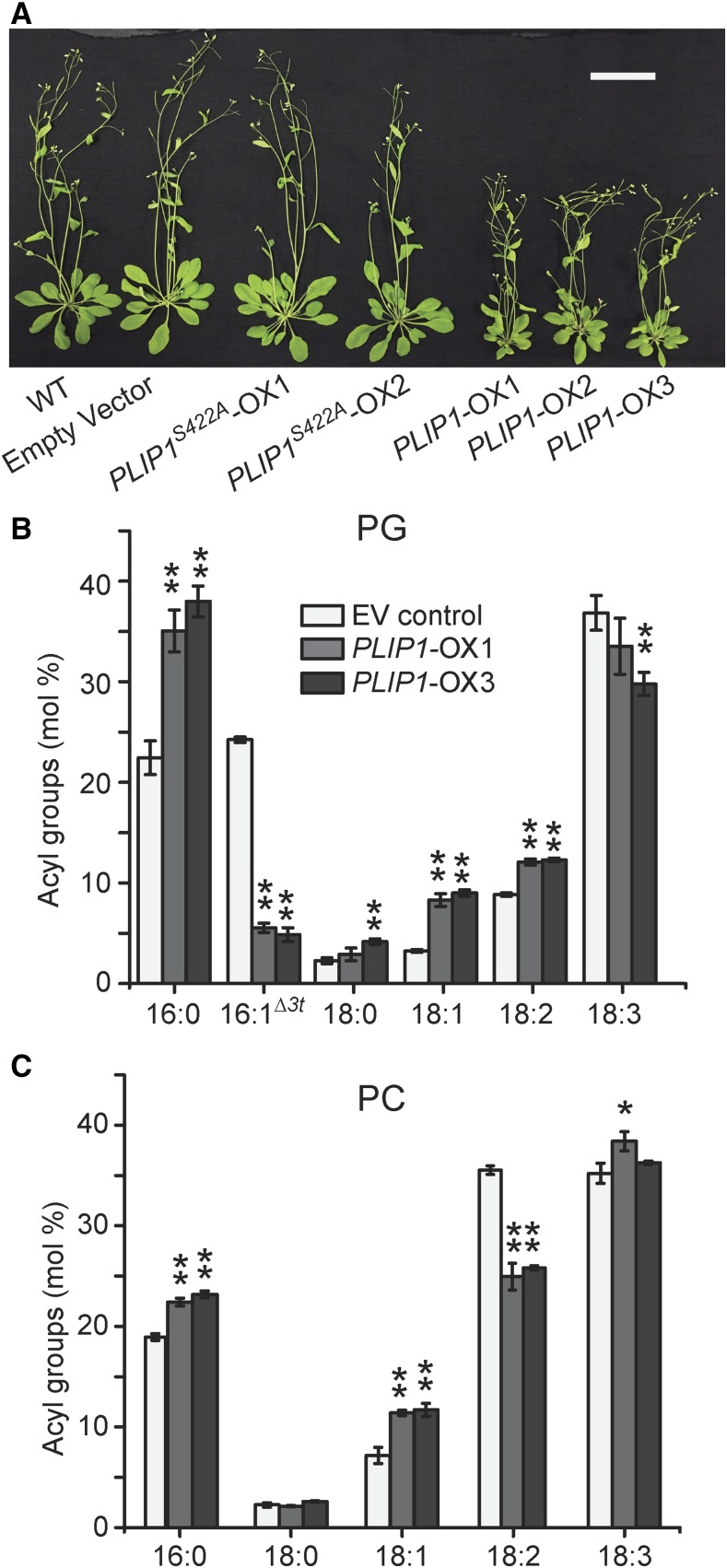
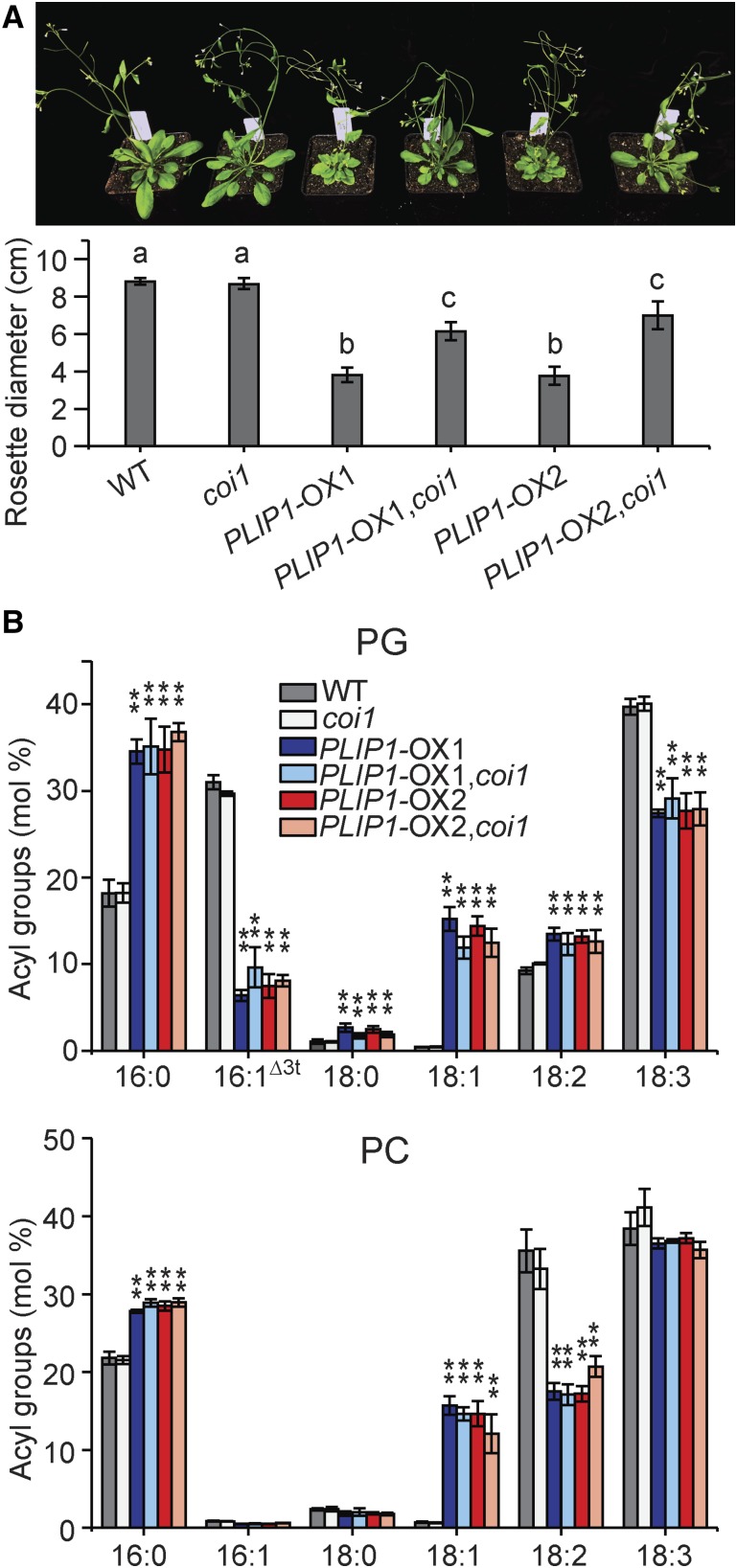
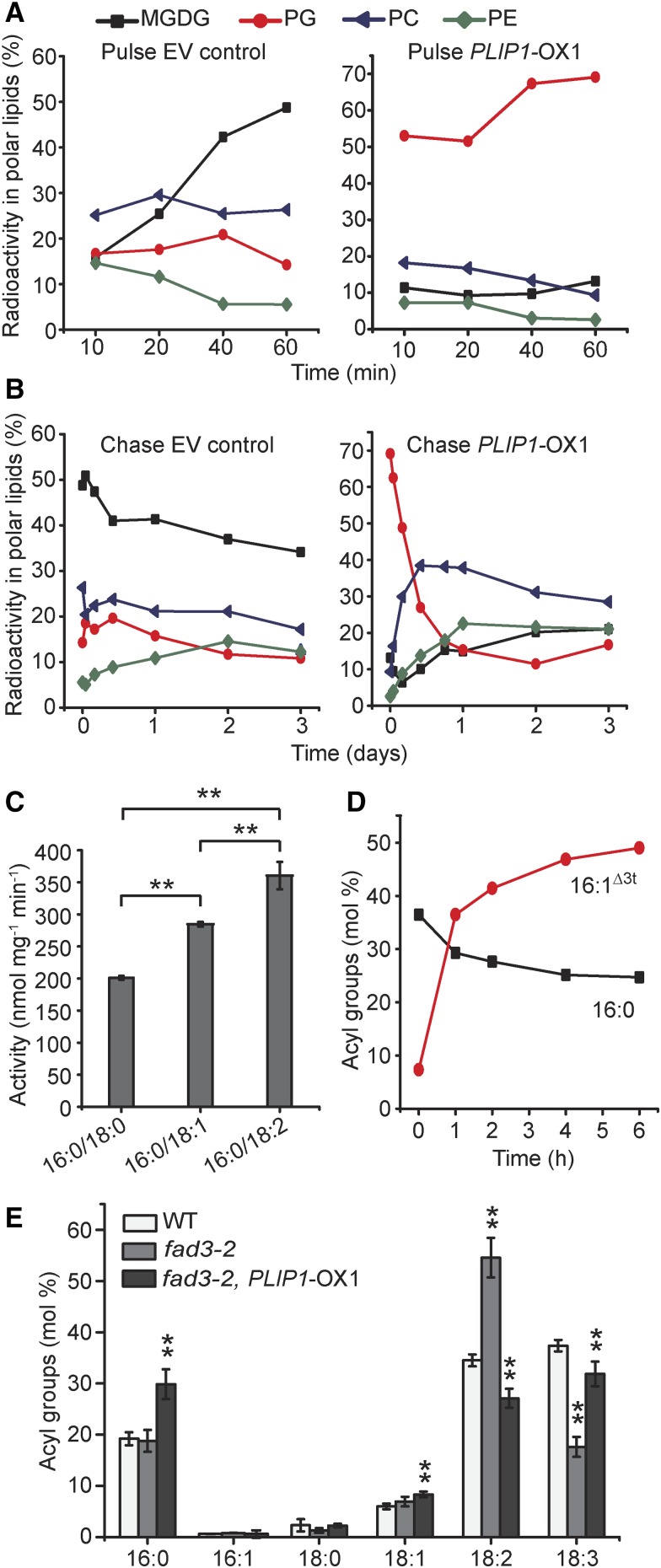
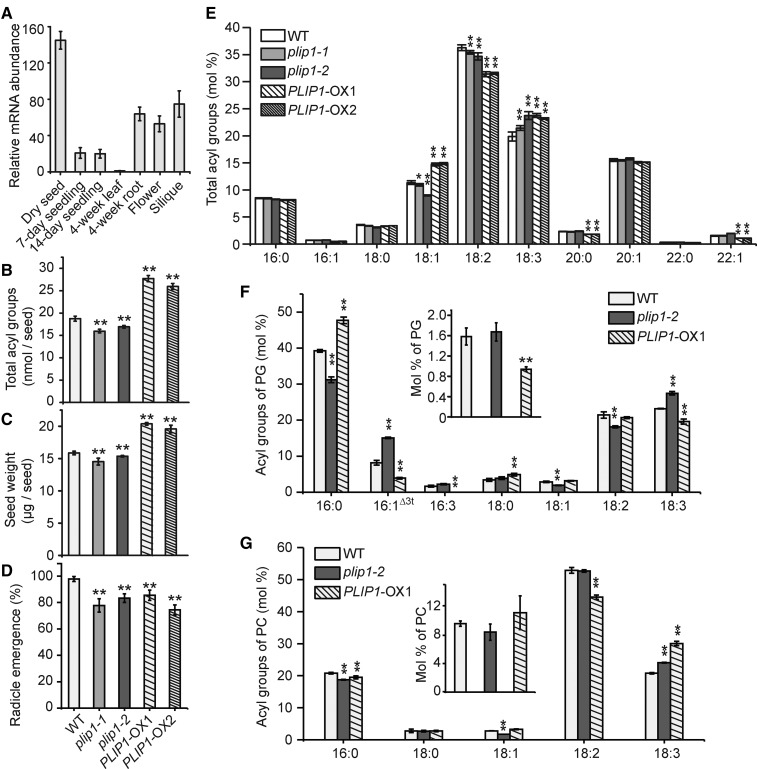
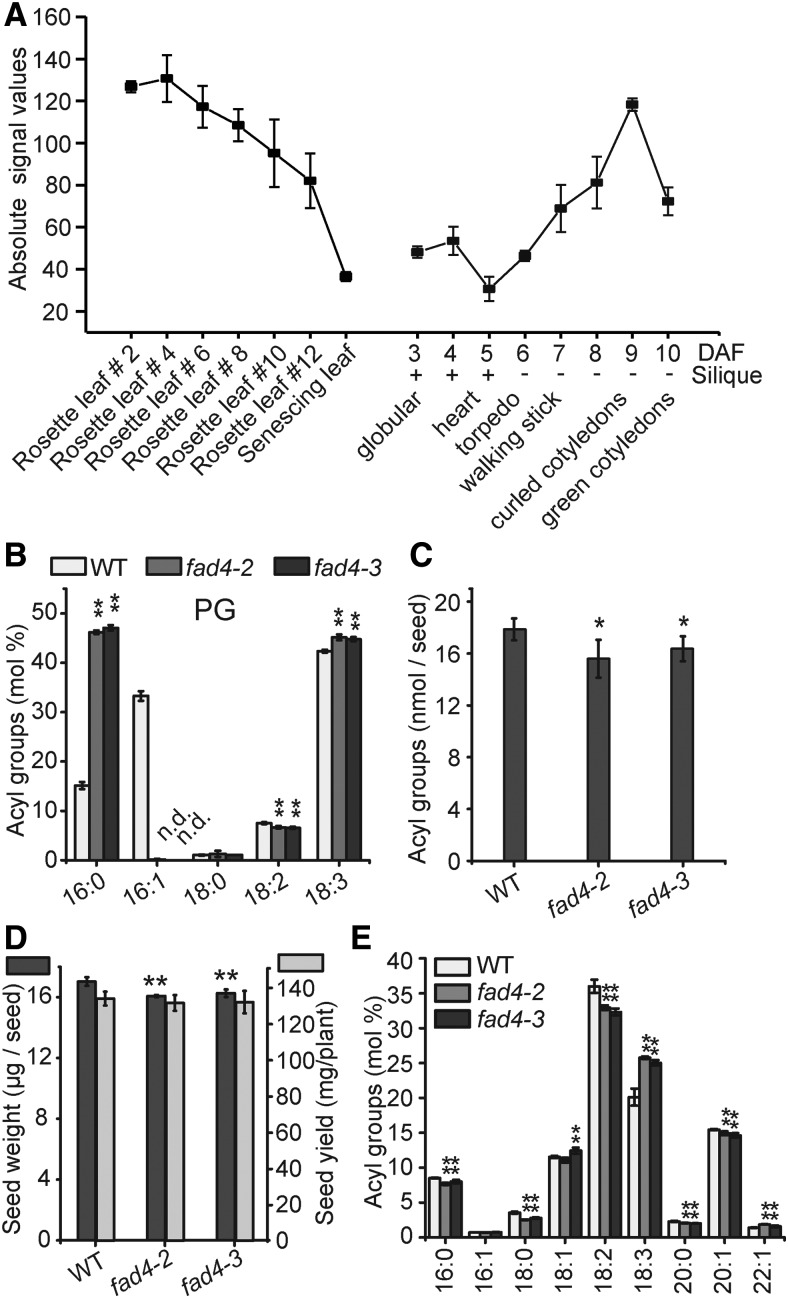
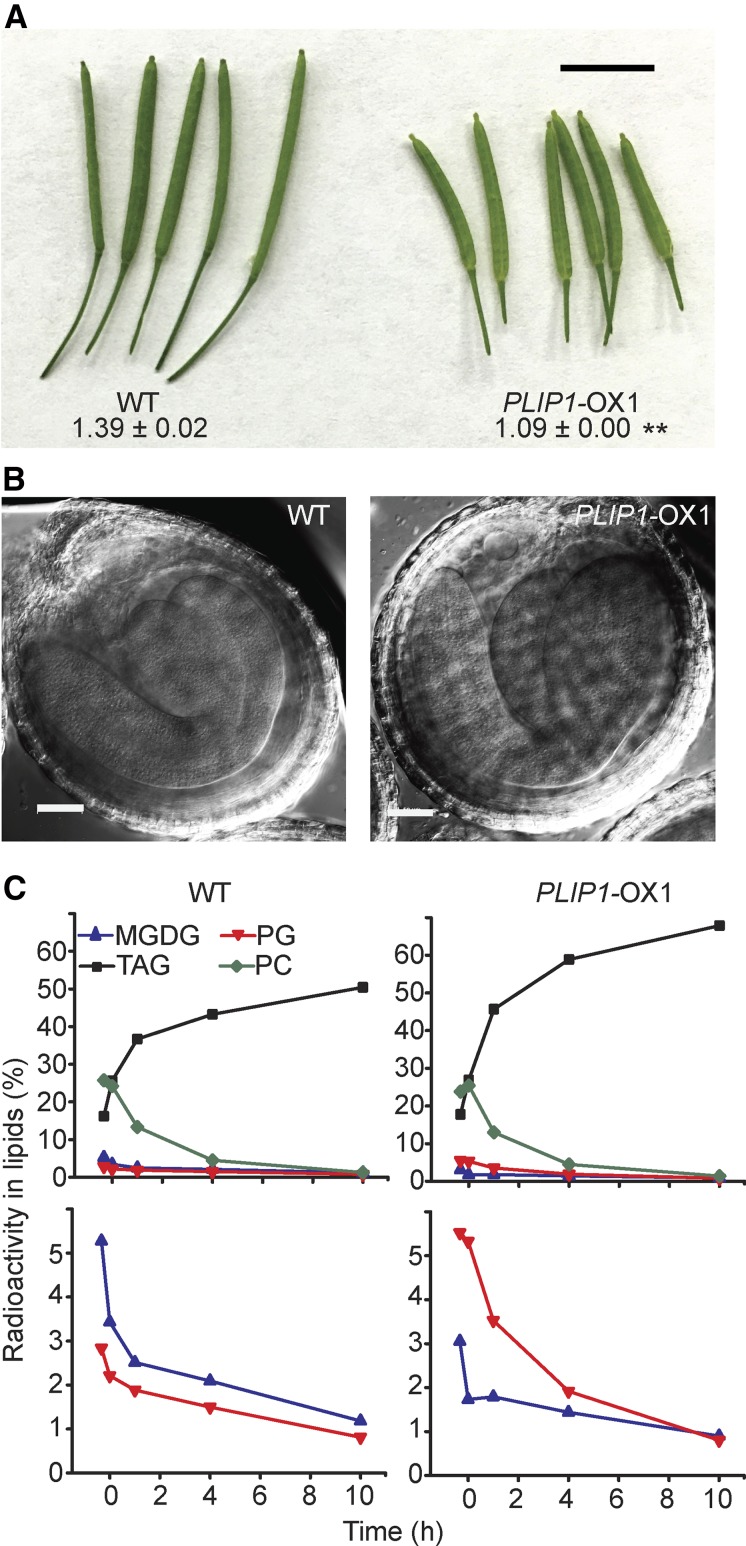
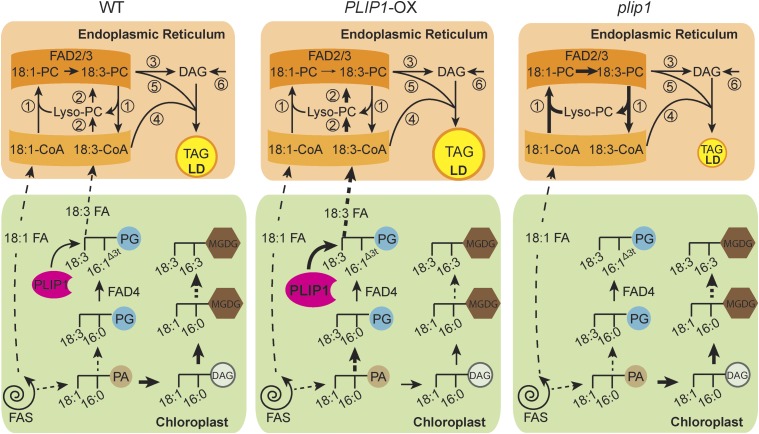
The lipid composition of thylakoid membranes inside chloroplasts is conserved from leaves to developing embryos. A finely tuned lipid assembly machinery is required to build these membranes during Arabidopsis thaliana development. Contrary to thylakoid lipid biosynthetic enzymes, the functions of most predicted chloroplast lipid-degrading enzymes remain to be elucidated. Here, we explore the biochemistry and physiological function of an Arabidopsis thylakoid membrane-associated lipase, PLASTID LIPASE1 (PLIP1). PLIP1 is a phospholipase A1 In vivo, PLIP1 hydrolyzes polyunsaturated acyl groups from a unique chloroplast-specific phosphatidylglycerol that contains 16:1 Δ3trans as its second acyl group. Thus far, a specific function of this 16:1 Δ3trans -containing phosphatidylglycerol in chloroplasts has remained elusive. The PLIP1 gene is highly expressed in seeds, and plip1 mutant seeds contain less oil and exhibit delayed germination compared with the wild type. Acyl groups released by PLIP1 are exported from the chloroplast, reincorporated into phosphatidylcholine, and ultimately enter seed triacylglycerol. Thus, 16:1 Δ3trans uniquely labels a small but biochemically active plastid phosphatidylglycerol pool in developing Arabidopsis embryos, which is subject to PLIP1 activity, thereby contributing a small fraction of the polyunsaturated fatty acids present in seed oil. We propose that acyl exchange involving thylakoid lipids functions in acyl export from plastids and seed oil biosynthesis.
© 2017 American Society of Plant Biologists. All rights reserved.
Figures
Comment in
-
Interconnecting plastid lipases.Nat Plants. 2018 May;4(5):240. doi: 10.1038/s41477-018-0154-5. Nat Plants. 2018. PMID: 29725090 No abstract available.
References
-
- Alonso J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657. - PubMed
-
- Andersson M.X., Dörmann P. (2009). Chloroplast membrane lipid biosynthesis and transport. In The Chloroplast, Sandelius A.S., Aronsson H., eds (Berlin, Heidelberg: Springer Berlin Heidelberg; ), pp. 125–158.
-
- Aronsson H., Jarvis P. (2002). A simple method for isolating import-competent Arabidopsis chloroplasts. FEBS Lett. 529: 215–220. - PubMed
-
- Bates P.D., Ohlrogge J.B., Pollard M. (2007). Incorporation of newly synthesized fatty acids into cytosolic glycerolipids in pea leaves occurs via acyl editing. J. Biol. Chem. 282: 31206–31216. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
