

Nuclear receptors connect progenitor transcription factors to cell cycle control
- PMID: 28687780
- PMCID: PMC5501803
- DOI: 10.1038/s41598-017-04936-7
Nuclear receptors connect progenitor transcription factors to cell cycle control
Abstract
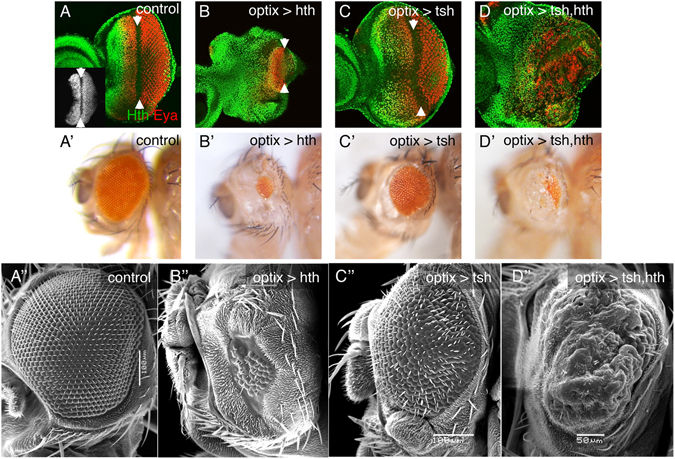
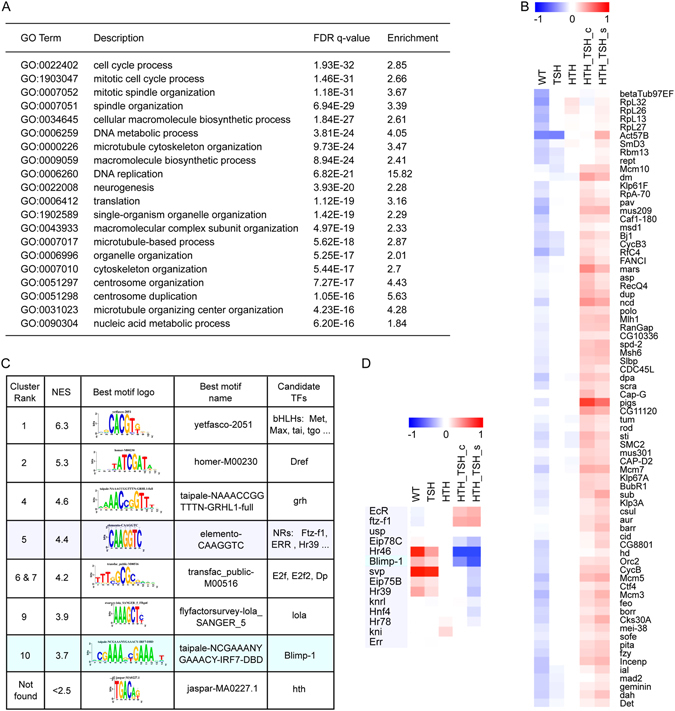
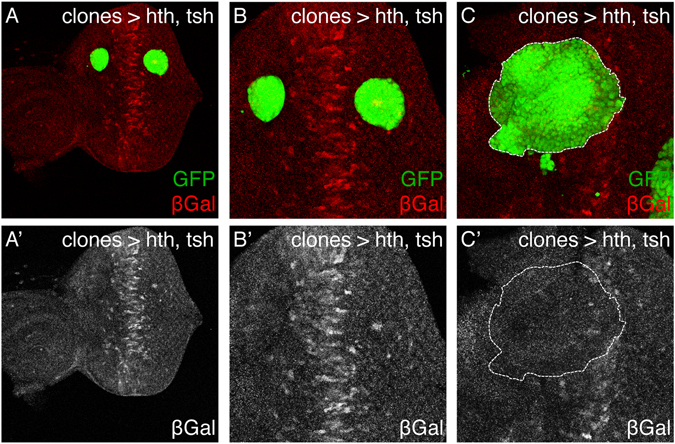
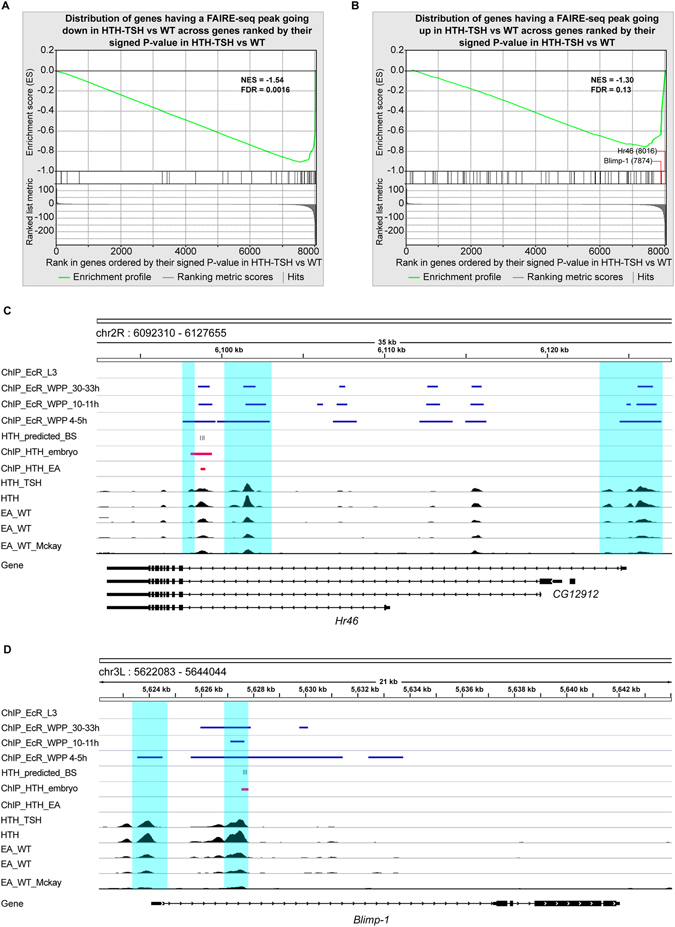
The specification and growth of organs is controlled simultaneously by networks of transcription factors. While the connection between these transcription factors with fate determinants is increasingly clear, how they establish the link with the cell cycle is far less understood. Here we investigate this link in the developing Drosophila eye, where two transcription factors, the MEIS1 homologue hth and the Zn-finger tsh, synergize to stimulate the proliferation of naïve eye progenitors. Experiments combining transcriptomics, open-chromatin profiling, motif analysis and functional assays indicate that these progenitor transcription factors exert a global regulation of the proliferation program. Rather than directly regulating cell cycle genes, they control proliferation through an intermediary layer of nuclear receptors of the ecdysone/estrogen-signaling pathway. This regulatory subnetwork between hth, tsh and nuclear receptors might be conserved from Drosophila to mammals, as we find a significant co-overexpression of their human homologues in specific cancer types.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Increased avidity for Dpp/BMP2 maintains the proliferation of progenitors-like cells in the Drosophila eye.Dev Biol. 2016 Oct 1;418(1):98-107. doi: 10.1016/j.ydbio.2016.08.004. Epub 2016 Aug 5. Dev Biol. 2016. PMID: 27502436
-
Differing strategies for the establishment and maintenance of teashirt and homothorax repression in the Drosophila wing.Development. 2004 Nov;131(22):5683-93. doi: 10.1242/dev.01450. Development. 2004. PMID: 15509768
-
Eye selector logic for a coordinated cell cycle exit.PLoS Genet. 2015 Feb 19;11(2):e1004981. doi: 10.1371/journal.pgen.1004981. eCollection 2015 Feb. PLoS Genet. 2015. PMID: 25695251 Free PMC article.
-
Ecdysone response gene E78 controls ovarian germline stem cell niche formation and follicle survival in Drosophila.Dev Biol. 2015 Apr 1;400(1):33-42. doi: 10.1016/j.ydbio.2015.01.013. Epub 2015 Jan 23. Dev Biol. 2015. PMID: 25624267 Free PMC article.
-
Atrophin proteins: an overview of a new class of nuclear receptor corepressors.Nucl Recept Signal. 2008;6:e009. doi: 10.1621/nrs.06009. Epub 2008 Oct 31. Nucl Recept Signal. 2008. PMID: 19043594 Free PMC article. Review.
Cited by
-
New regulators of Drosophila eye development identified from temporal transcriptome changes.Genetics. 2021 Apr 15;217(4):iyab007. doi: 10.1093/genetics/iyab007. Genetics. 2021. PMID: 33681970 Free PMC article.
-
Combined inactivation of RB and Hippo converts differentiating Drosophila photoreceptors into eye progenitor cells through derepression of homothorax.Dev Cell. 2023 Nov 6;58(21):2261-2274.e6. doi: 10.1016/j.devcel.2023.09.003. Epub 2023 Oct 16. Dev Cell. 2023. PMID: 37848027 Free PMC article.
-
A single-cell transcriptomic atlas characterizes the silk-producing organ in the silkworm.Nat Commun. 2022 Jun 9;13(1):3316. doi: 10.1038/s41467-022-31003-1. Nat Commun. 2022. PMID: 35680954 Free PMC article.
-
Variation in Pleiotropic Hub Gene Expression Is Associated with Interspecific Differences in Head Shape and Eye Size in Drosophila.Mol Biol Evol. 2021 May 4;38(5):1924-1942. doi: 10.1093/molbev/msaa335. Mol Biol Evol. 2021. PMID: 33386848 Free PMC article.
-
Feedback control of organ size precision is mediated by BMP2-regulated apoptosis in the Drosophila eye.PLoS Biol. 2024 Jan 30;22(1):e3002450. doi: 10.1371/journal.pbio.3002450. eCollection 2024 Jan. PLoS Biol. 2024. PMID: 38289899 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
