

VAMP8-mediated NOX2 recruitment to endosomes is necessary for antigen release
- PMID: 28688576
- PMCID: PMC5641923
- DOI: 10.1016/j.ejcb.2017.06.007
VAMP8-mediated NOX2 recruitment to endosomes is necessary for antigen release
Abstract
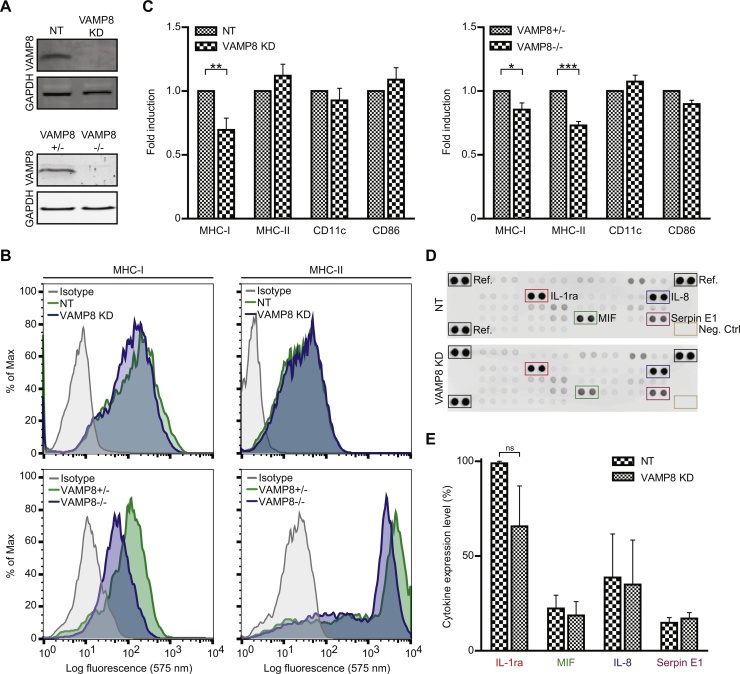
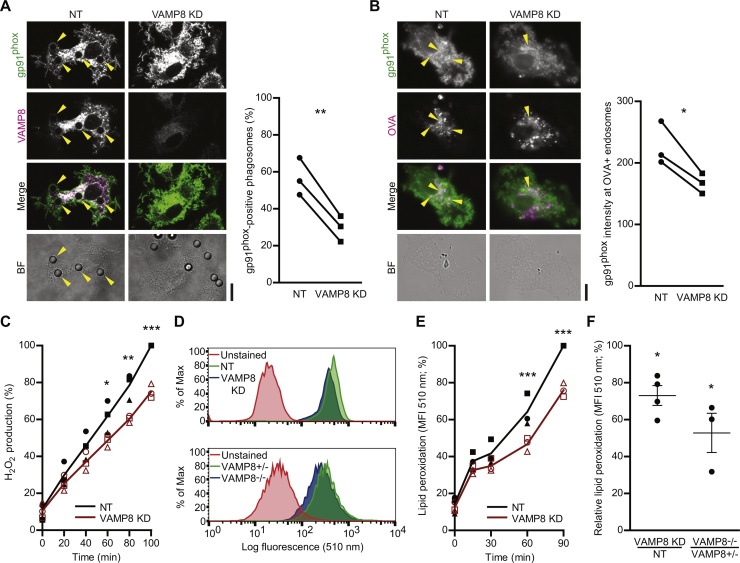
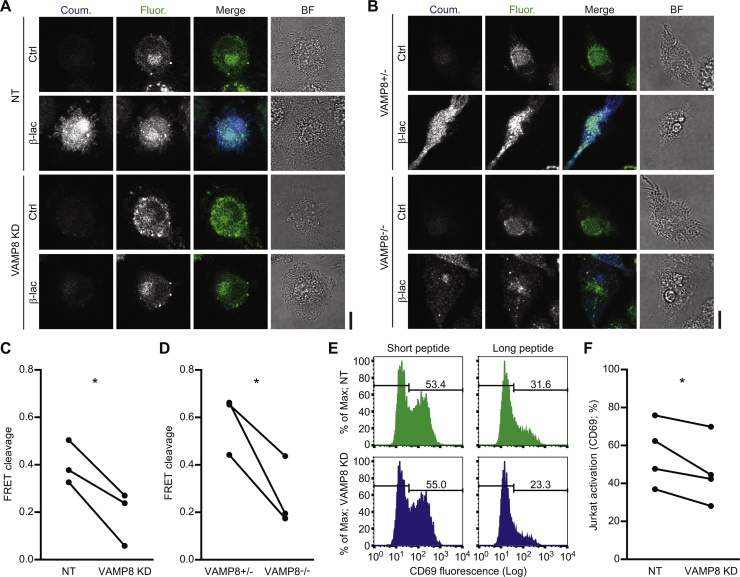
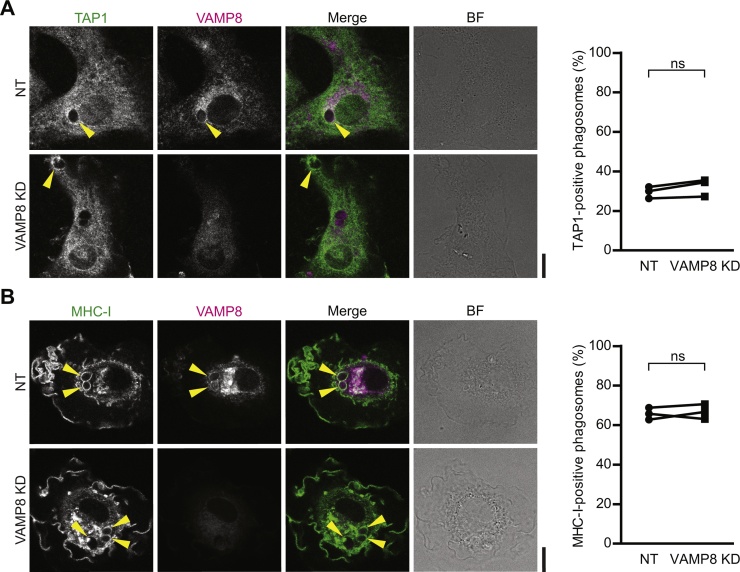
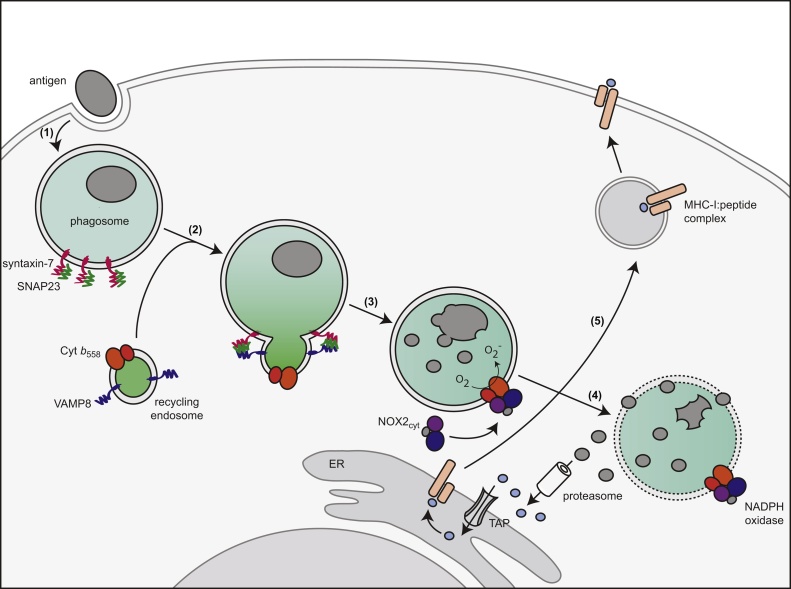
Cross-presentation of foreign antigen in major histocompatibility complex (MHC) class I by dendritic cells (DCs) requires activation of the NADPH-oxidase NOX2 complex. We recently showed that NOX2 is recruited to phagosomes by the SNARE protein VAMP8 where NOX2-produced reactive oxygen species (ROS) cause lipid oxidation and membrane disruption, promoting antigen translocation into the cytosol for cross-presentation. In this study, we extend these findings by showing that VAMP8 is also involved in NOX2 trafficking to endosomes. Moreover, we demonstrate in both human and mouse DCs that absence of VAMP8 leads to decreased ROS production, lipid peroxidation and antigen translocation, and that this impairs cross-presentation. In contrast, knockdown of VAMP8 did not affect recruitment of MHC class I and the transporter associated with antigen processing 1 (TAP1) to phagosomes, although surface levels of MHC class I were reduced. Thus, in addition to a secretory role, VAMP8-mediates trafficking of NOX2 to endosomes and phagosomes and this promotes induction of cytolytic T cell immune responses.
Keywords: Cross-presentation; Dendritic cells; Lipid peroxidation; NOX2; VAMP8.
Copyright © 2017 The Authors. Published by Elsevier GmbH.. All rights reserved.
Figures
References
-
- Allan E.R.O., Tailor P., Balce D.R., Pirzadeh P., McKenna N.T., Renaux B., Warren A.L., Jirik F.R., Yates R.M. NADPH oxidase modifies patterns of MHC class II-restricted epitopic repertoires through redox control of antigen processing. J. Immunol. 2014;192:4989–5001. - PubMed
-
- Arango Duque G., Fukuda M., Descoteaux A. Synaptotagmin XI regulates phagocytosis and cytokine secretion in macrophages. J. Immunol. 2013;190:1737–1745. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
