

The role of succinate and ROS in reperfusion injury - A critical appraisal
- PMID: 28689004
- PMCID: PMC5678286
- DOI: 10.1016/j.yjmcc.2017.06.016
The role of succinate and ROS in reperfusion injury - A critical appraisal
Abstract
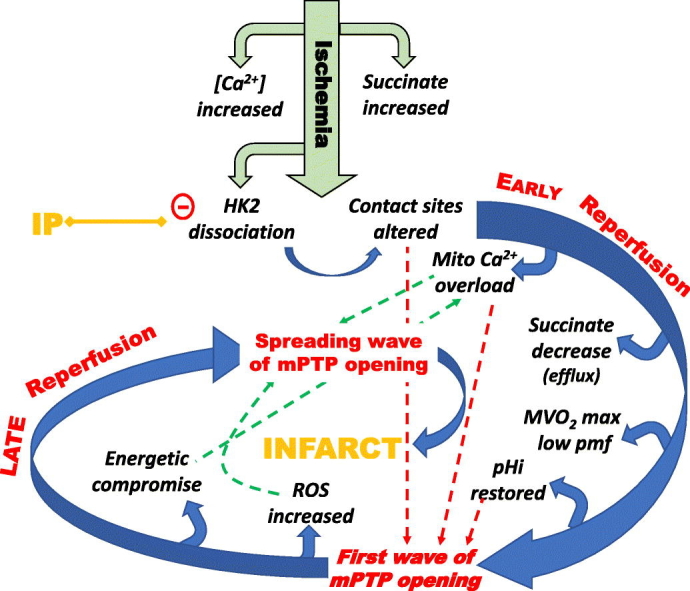
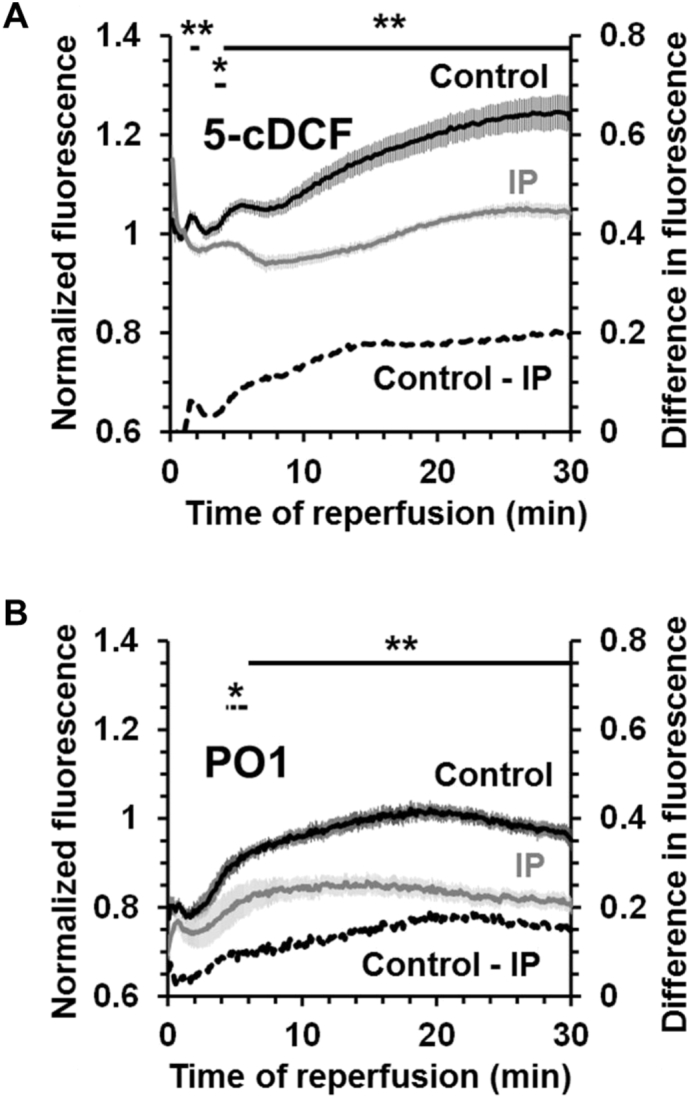
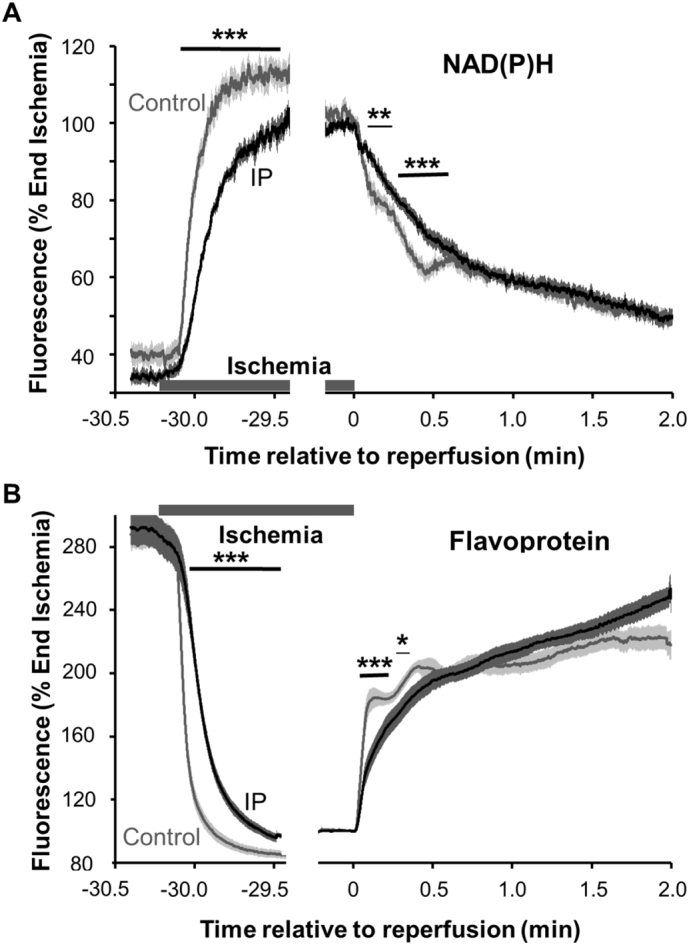
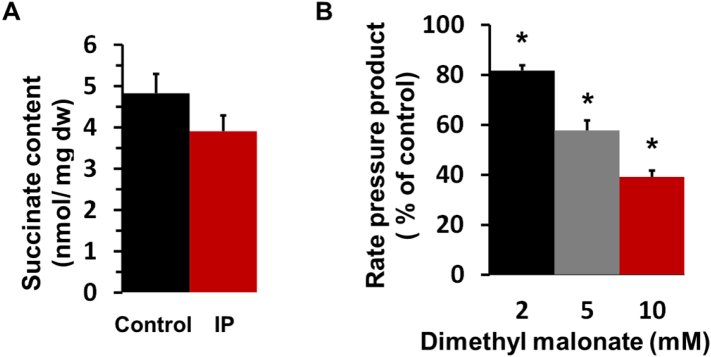
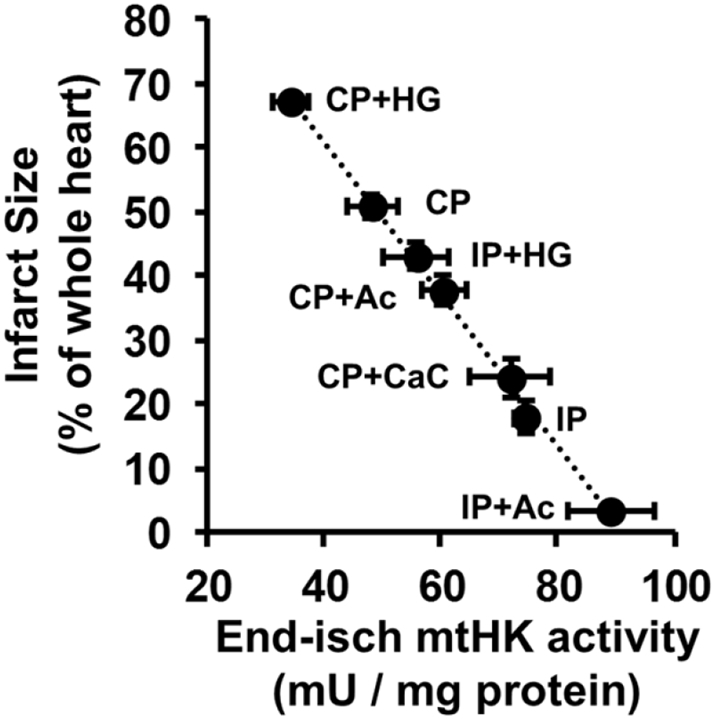
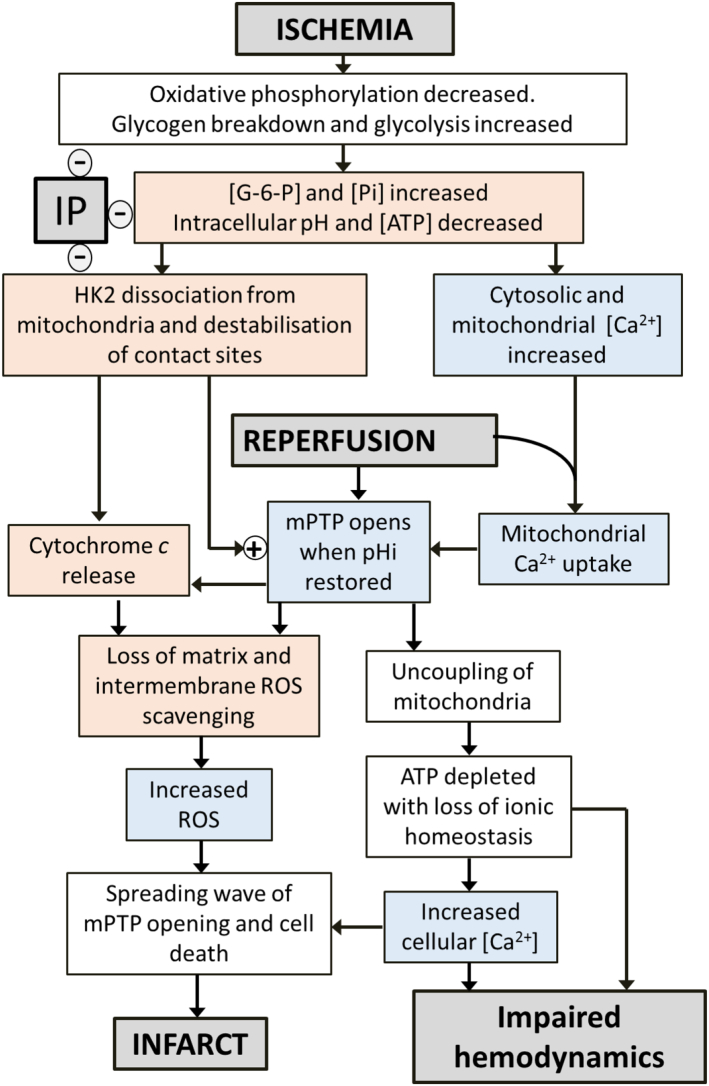
We critically assess the proposal that succinate-fuelled reverse electron flow (REF) drives mitochondrial matrix superoxide production from Complex I early in reperfusion, thus acting as a key mediator of ischemia/reperfusion (IR) injury. Real-time surface fluorescence measurements of NAD(P)H and flavoprotein redox state suggest that conditions are unfavourable for REF during early reperfusion. Furthermore, rapid loss of succinate accumulated during ischemia can be explained by its efflux rather than oxidation. Moreover, succinate accumulation during ischemia is not attenuated by ischemic preconditioning (IP) despite powerful cardioprotection. In addition, measurement of intracellular reactive oxygen species (ROS) during reperfusion using surface fluorescence and mitochondrial aconitase activity detected major increases in ROS only after mitochondrial permeability transition pore (mPTP) opening was first detected. We conclude that mPTP opening is probably triggered initially by factors other than ROS, including increased mitochondrial [Ca2+]. However, IP only attenuates [Ca2+] increases later in reperfusion, again after initial mPTP opening, implying that IP regulates mPTP opening through additional mechanisms. One such is mitochondria-bound hexokinase 2 (HK2) which dissociates from mitochondria during ischemia in control hearts but not those subject to IP. Indeed, there is a strong correlation between the extent of HK2 loss from mitochondria during ischemia and infarct size on subsequent reperfusion. Mechanisms linking HK2 dissociation to mPTP sensitisation remain to be fully established but several related processes have been implicated including VDAC1 oligomerisation, the stability of contact sites between the inner and outer membranes, cristae morphology, Bcl-2 family members and mitochondrial fission proteins such as Drp1.
Keywords: Calcium; Heart; Hexokinase; Ischemia/reperfusion injury; Mitochondria; Permeability transition pore; Reactive oxygen species; Succinate.
Copyright © 2017 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
Comment in
-
Letter in response to "the role of succinate and ROS in reperfusion injury - A critical appraisal" by Andrienko et al.J Mol Cell Cardiol. 2017 Nov;112:131. doi: 10.1016/j.yjmcc.2017.08.010. Epub 2017 Aug 24. J Mol Cell Cardiol. 2017. PMID: 28842242 No abstract available.
References
-
- Halestrap A.P. A pore way to die: the role of mitochondria in reperfusion injury and cardioprotection. Biochem. Soc. Trans. 2010;38:841–860. - PubMed
-
- Halestrap A.P., Connern C.P., Griffiths E.J., Kerr P.M. Cyclosporin A binding to mitochondrial cyclophilin inhibits the permeability transition pore and protects hearts from ischaemia/reperfusion injury. Mol. Cell. Biochem. 1997;174:167–172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
