

The effects of historical fragmentation on major histocompatibility complex class II β and microsatellite variation in the Aegean island reptile, Podarcis erhardii
- PMID: 28690787
- PMCID: PMC5496512
- DOI: 10.1002/ece3.3022
The effects of historical fragmentation on major histocompatibility complex class II β and microsatellite variation in the Aegean island reptile, Podarcis erhardii
Abstract
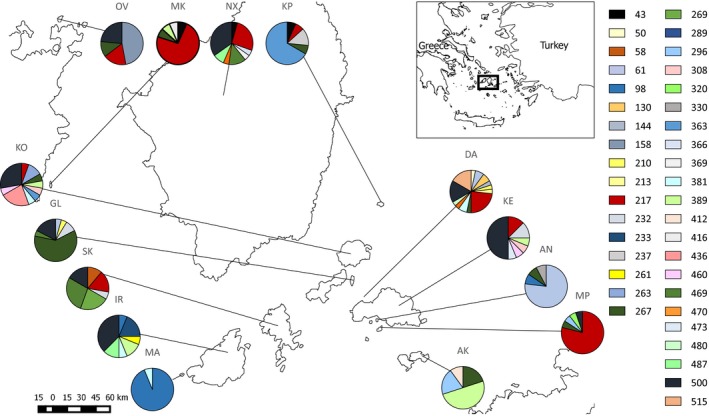
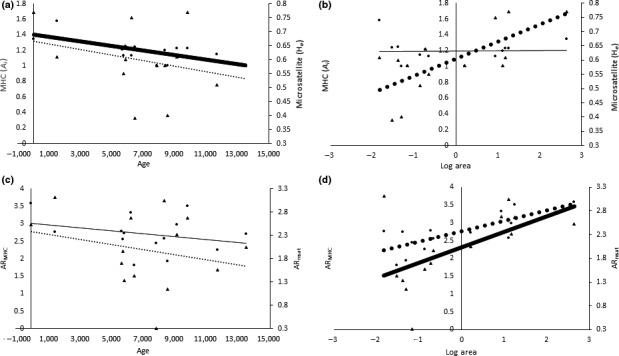
The major histocompatibility complex (MHC) plays a key role in disease resistance and is the most polymorphic gene region in vertebrates. Although habitat fragmentation is predicted to lead to a loss in MHC variation through drift, the impact of other evolutionary forces may counter this effect. Here we assess the impact of selection, drift, migration, and recombination on MHC class II and microsatellite variability in 14 island populations of the Aegean wall lizard Podarcis erhardii. Lizards were sampled from islands within the Cyclades (Greece) formed by rising sea levels as the last glacial maximum approximately 20,000 before present. Bathymetric data were used to determine the area and age of each island, allowing us to infer the corresponding magnitude and timing of genetic bottlenecks associated with island formation. Both MHC and microsatellite variation were positively associated with island area, supporting the hypothesis that drift governs neutral and adaptive variation in this system. However, MHC but not microsatellite variability declined significantly with island age. This discrepancy is likely due to the fact that microsatellites attain mutation-drift equilibrium more rapidly than MHC. Although we detected signals of balancing selection, recombination and migration, the effects of these evolutionary processes appeared negligible relative to drift. This study demonstrates how land bridge islands can provide novel insights into the impact of historical fragmentation on genetic diversity as well as help disentangle the effects of different evolutionary forces on neutral and adaptive diversity.
Keywords: drift; historical fragmentation; immunity; major histocompatibility complex; selection.
Figures




References
-
- Altizer, S. , Harvell, D. , & Friedle, E. (2003). Rapid evolutionary dynamics and disease threats to biodiversity. Trends in Ecology and Evolution, 18, 589–596.
-
- Altschul, S. , Gish, W. , Miller, W. , Myers, E. , & Lipman, D. (1990). Basic local alignment search tool. Journal of Molecular Biology, 215, 403–410. - PubMed
-
- Babik, W. , Taberlet, P. , Ejsmond, M. J. , & Radwan, J. (2009). New generation sequencers as a tool for genotyping of highly polymorphic multilocus MHC system. Molecular Ecology Resources, 9, 713–719. - PubMed
-
- Bernatchez, L. , & Landry, C. (2003). MHC studies in nonmodel vertebrates: What have we learned about natural selection in 15 years? Journal of Evolutionary Biology, 16, 363–377. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
