

Site-Specific Phosphorylation of PSD-95 PDZ Domains Reveals Fine-Tuned Regulation of Protein-Protein Interactions
- PMID: 28692247
- PMCID: PMC6081957
- DOI: 10.1021/acschembio.7b00361
Site-Specific Phosphorylation of PSD-95 PDZ Domains Reveals Fine-Tuned Regulation of Protein-Protein Interactions
Abstract
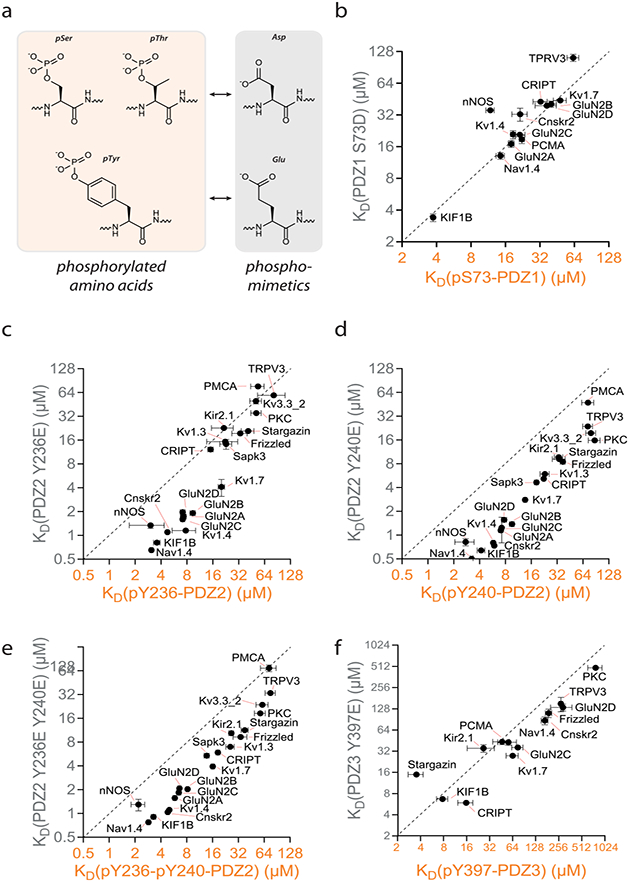
The postsynaptic density protein of 95 kDa (PSD-95) is a key scaffolding protein that controls signaling at synapses in the brain through interactions of its PDZ domains with the C-termini of receptors, ion channels, and enzymes. PSD-95 is highly regulated by phosphorylation. To explore the effect of phosphorylation on PSD-95, we used semisynthetic strategies to introduce phosphorylated amino acids at four positions within the PDZ domains and examined the effects on interactions with a large set of binding partners. We observed complex effects on affinity. Most notably, phosphorylation at Y397 induced a significant increase in affinity for stargazin, as confirmed by NMR and single molecule FRET. Additionally, we compared the effects of phosphorylation to phosphomimetic mutations, which revealed that phosphomimetics are ineffective substitutes for tyrosine phosphorylation. Our strategy to generate site-specifically phosphorylated PDZ domains provides a detailed understanding of the role of phosphorylation in the regulation of PSD-95 interactions.
Conflict of interest statement
The authors declare no competing financial interest.
Figures






References
-
- Sheng M, and Sala C (2001) PDZ domains and the organization of supramolecular complexes. Annu. Rev. Neurosci 24, 1–29. - PubMed
-
- Kornau HC, Schenker LT, Kennedy MB, and Seeburg PH (1995) Domain interaction between NMDA receptor subunits and the postsynaptic density protein PSD-95. Science 269, 1737–1740. - PubMed
-
- Feng W, and Zhang M (2009) Organization and dynamics of PDZ-domain-related supramodules in the postsynaptic density. Nat. Rev. Neurosci 10, 87–99. - PubMed
-
- Oliva C, Escobedo P, Astorga C, Molina C, and Sierralta J (2012) Role of the MAGUK protein family in synapse formation and function. Dev. Neurobiol 72, 57–72. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
