

The contribution of the C5 protein subunit of Escherichia coli ribonuclease P to specificity for precursor tRNA is modulated by proximal 5' leader sequences
- PMID: 28694328
- PMCID: PMC5602109
- DOI: 10.1261/rna.056408.116
The contribution of the C5 protein subunit of Escherichia coli ribonuclease P to specificity for precursor tRNA is modulated by proximal 5' leader sequences
Abstract
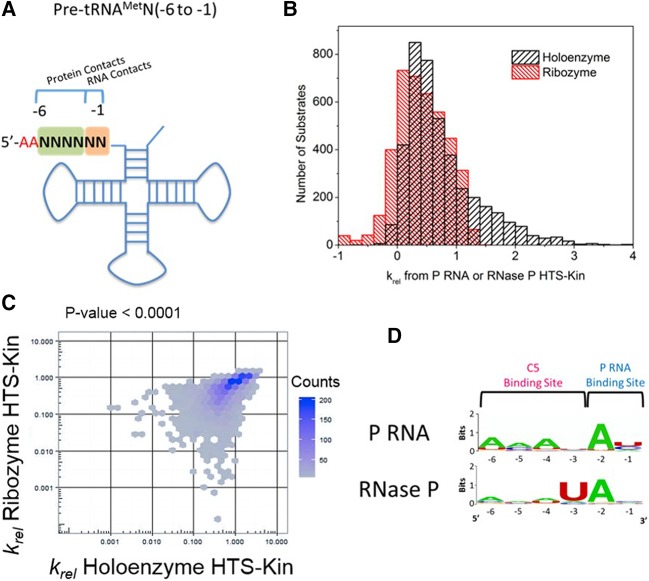
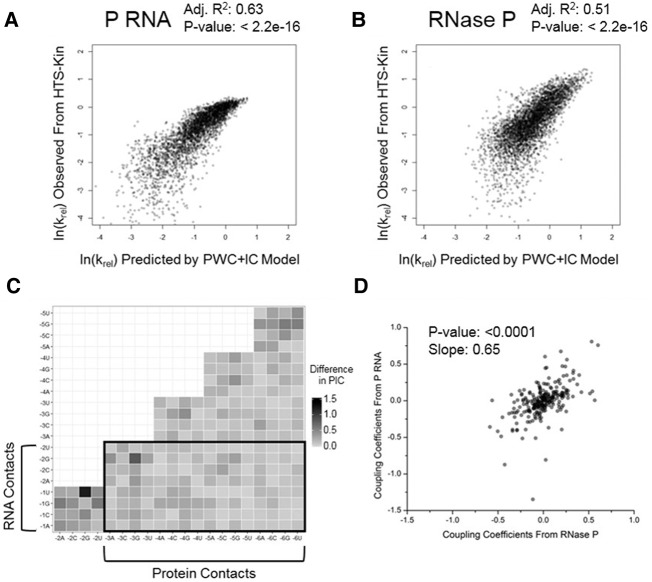
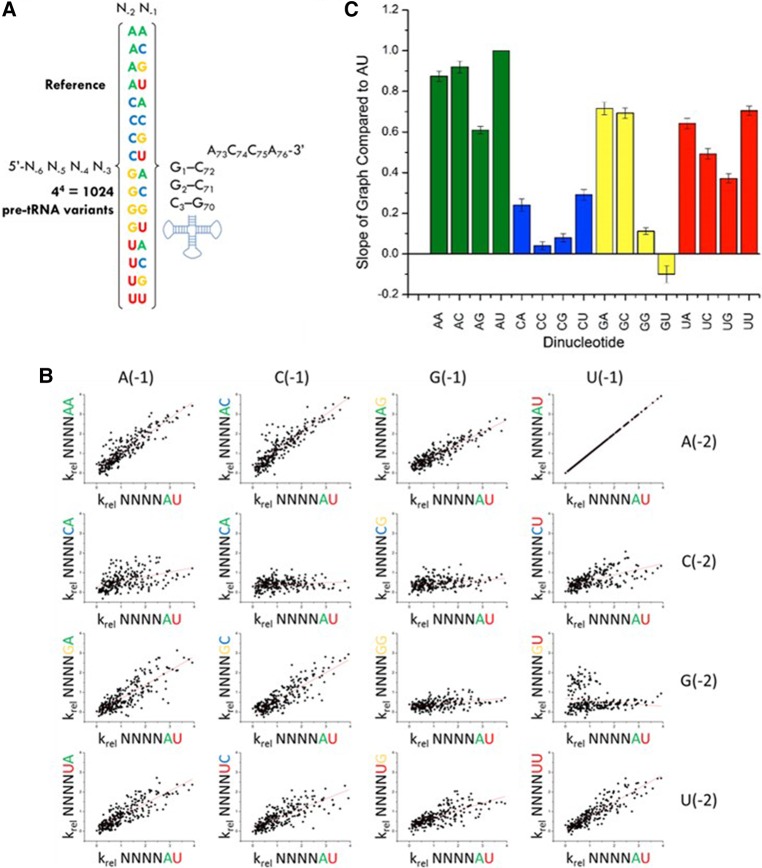
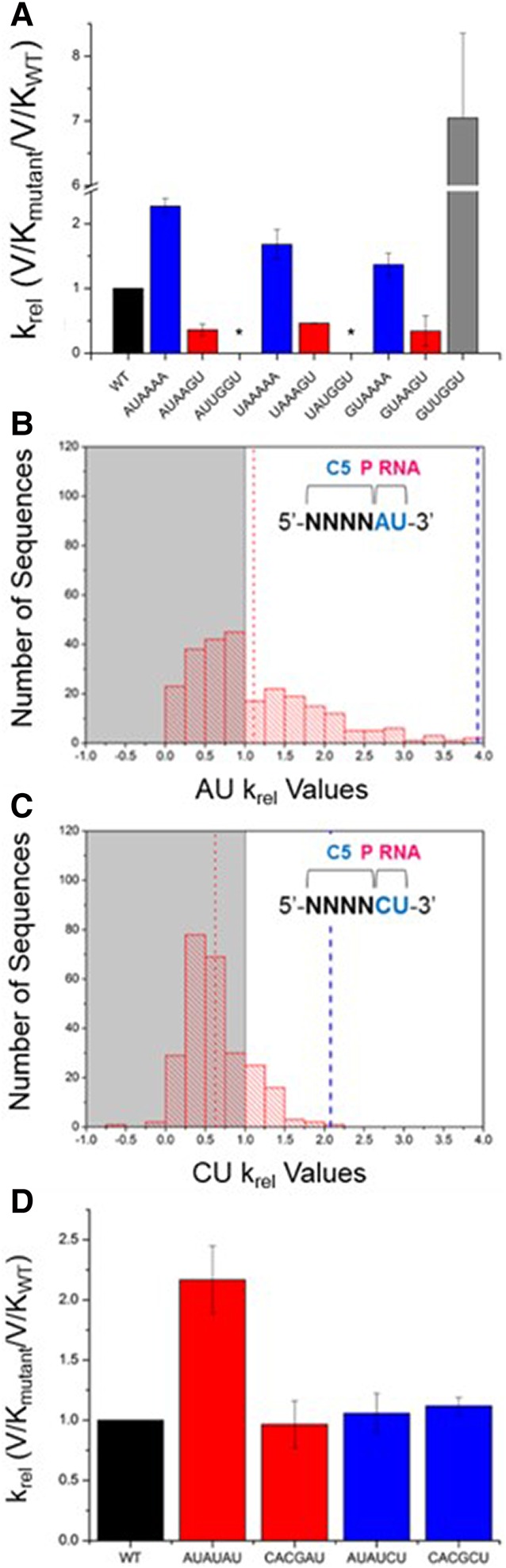
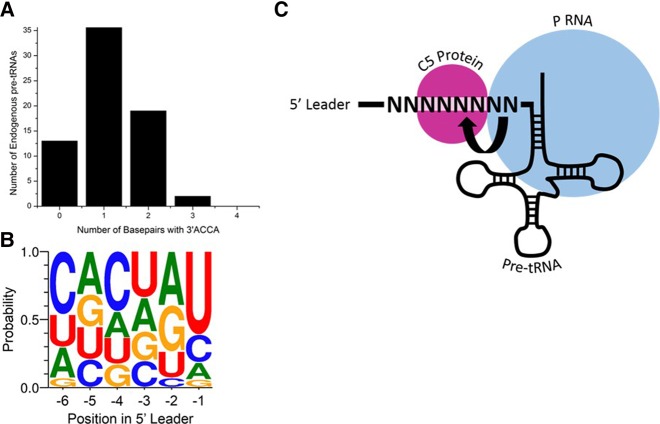
Recognition of RNA by RNA processing enzymes and RNA binding proteins often involves cooperation between multiple subunits. However, the interdependent contributions of RNA and protein subunits to molecular recognition by ribonucleoproteins are relatively unexplored. RNase P is an endonuclease that removes 5' leaders from precursor tRNAs and functions in bacteria as a dimer formed by a catalytic RNA subunit (P RNA) and a protein subunit (C5 in E. coli). The P RNA subunit contacts the tRNA body and proximal 5' leader sequences [N(-1) and N(-2)] while C5 binds distal 5' leader sequences [N(-3) to N(-6)]. To determine whether the contacts formed by P RNA and C5 contribute independently to specificity or exhibit cooperativity or anti-cooperativity, we compared the relative kcat/Km values for all possible combinations of the six proximal 5' leader nucleotides (n = 4096) for processing by the E. coli P RNA subunit alone and by the RNase P holoenzyme. We observed that while the P RNA subunit shows specificity for 5' leader nucleotides N(-2) and N(-1), the presence of the C5 protein reduces the contribution of P RNA to specificity, but changes specificity at N(-2) and N(-3). The results reveal that the contribution of C5 protein to RNase P processing is controlled by the identity of N(-2) in the pre-tRNA 5' leader. The data also clearly show that pairing of the 5' leader with the 3' ACCA of tRNA acts as an anti-determinant for RNase P cleavage. Comparative analysis of genomically encoded E. coli tRNAs reveals that both anti-determinants are subject to negative selection in vivo.
Keywords: RNase P; enzyme specificity; high-throughput sequencing; kinetics; molecular recognition; pre-tRNA processing.
© 2017 Niland et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
-
- Altman S, Guerrier-Takada C. 1986. M1 RNA, the RNA subunit of Escherichia coli ribonuclease P, can undergo a pH-sensitive conformational change. Biochemistry 25: 1205–1208. - PubMed
-
- Anderson VE. 2015. Multiple alternative substrate kinetics. Biochim Biophys Acta 1854: 1729–1736. - PubMed
-
- Brännvall M, Mattsson JG, Svärd SG, Kirsebom LA. 1998. RNase P RNA structure and cleavage reflect the primary structure of tRNA genes. J Mol Biol 283: 771–783. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous