

Monocular deprivation induces dendritic spine elimination in the developing mouse visual cortex
- PMID: 28694464
- PMCID: PMC5504056
- DOI: 10.1038/s41598-017-05337-6
Monocular deprivation induces dendritic spine elimination in the developing mouse visual cortex
Abstract
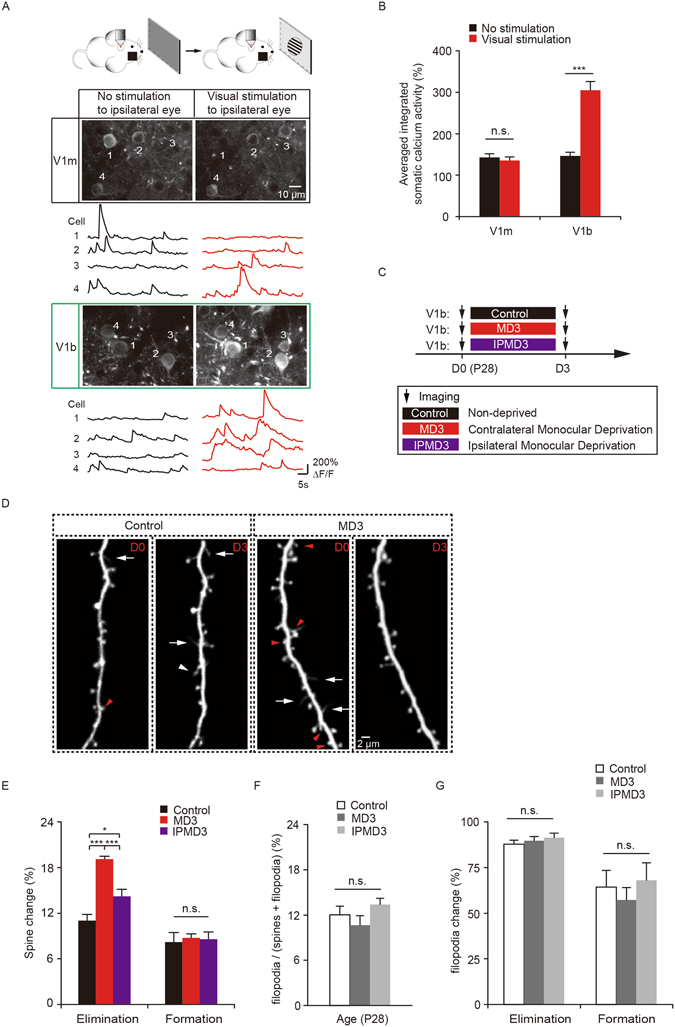
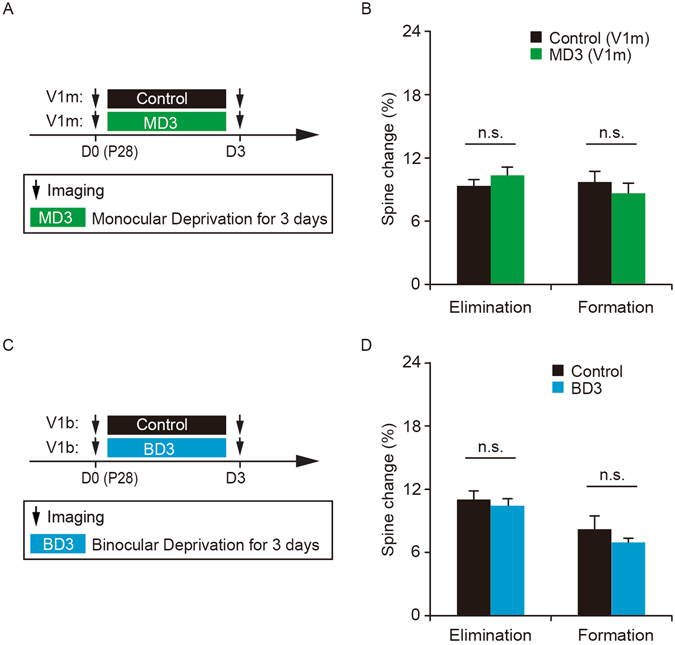
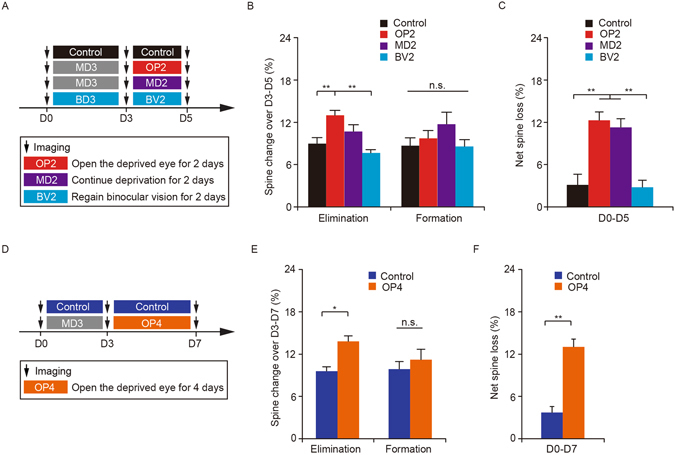
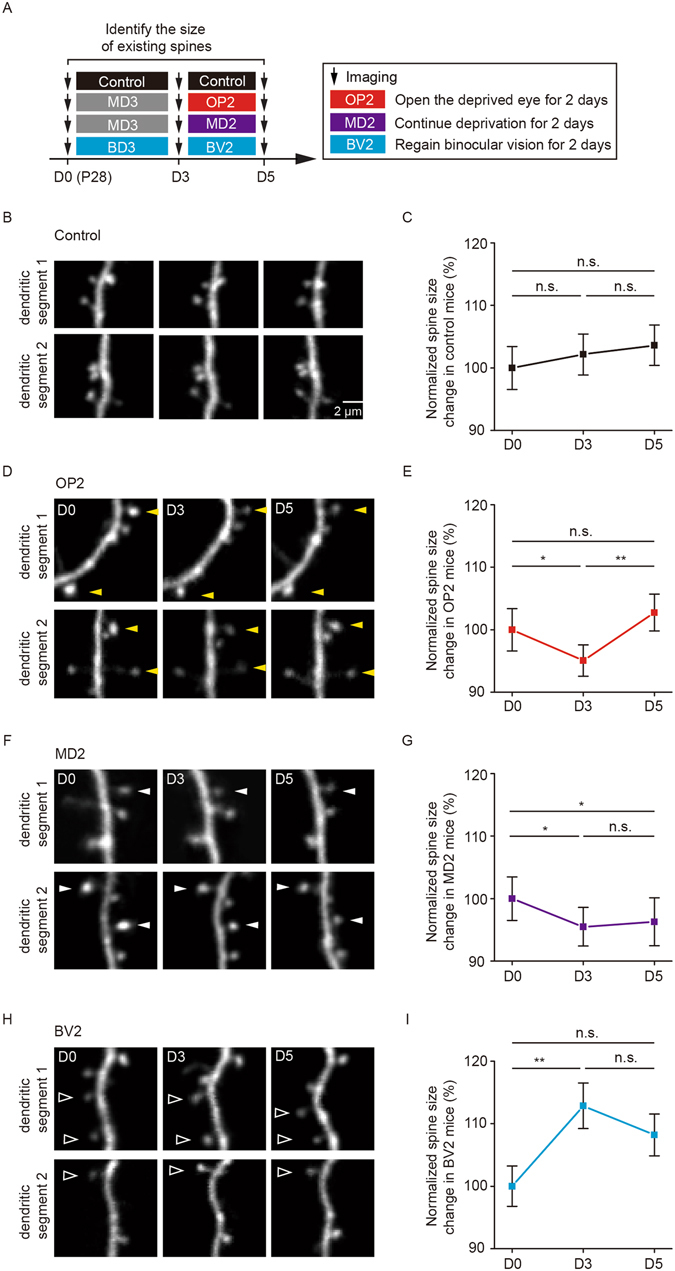
It is well established that visual deprivation has a profound impact on the responsiveness of neurons in the developing visual cortex. The effect of visual deprivation on synaptic connectivity remains unclear. Using transcranial two-photon microscopy, we examined the effect of visual deprivation and subsequent recovery on dendritic spine remodeling of layer 5 pyramidal neurons in the mouse primary visual cortex. We found that monocular deprivation (MD), but not binocular deprivation (BD), increased dendritic spine elimination over 3 days in the binocular region of 4-week-old adolescent mice. This MD-induced dendritic spine elimination persisted during subsequent 2-4 days of binocular recovery. Furthermore, we found that average dendritic spine sizes were decreased and increased following 3-day MD and BD, respectively. These spine size changes induced by MD or BD tended to be reversed during subsequent binocular recovery. Taken together, these findings reveal differential effects of MD and BD on synaptic connectivity of layer 5 pyramidal neurons and underscore the persistent impact of MD on synapse loss in the developing visual cortex.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Hubel DH, Wiesel TN. Binocular interaction in striate cortex of Kittens reared with artificial squint. Journal of Neurophysiology. 1965;28:1041–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
