

Amelogenesis Imperfecta; Genes, Proteins, and Pathways
- PMID: 28694781
- PMCID: PMC5483479
- DOI: 10.3389/fphys.2017.00435
Amelogenesis Imperfecta; Genes, Proteins, and Pathways
Abstract
Amelogenesis imperfecta (AI) is the name given to a heterogeneous group of conditions characterized by inherited developmental enamel defects. AI enamel is abnormally thin, soft, fragile, pitted and/or badly discolored, with poor function and aesthetics, causing patients problems such as early tooth loss, severe embarrassment, eating difficulties, and pain. It was first described separately from diseases of dentine nearly 80 years ago, but the underlying genetic and mechanistic basis of the condition is only now coming to light. Mutations in the gene AMELX, encoding an extracellular matrix protein secreted by ameloblasts during enamel formation, were first identified as a cause of AI in 1991. Since then, mutations in at least eighteen genes have been shown to cause AI presenting in isolation of other health problems, with many more implicated in syndromic AI. Some of the encoded proteins have well documented roles in amelogenesis, acting as enamel matrix proteins or the proteases that degrade them, cell adhesion molecules or regulators of calcium homeostasis. However, for others, function is less clear and further research is needed to understand the pathways and processes essential for the development of healthy enamel. Here, we review the genes and mutations underlying AI presenting in isolation of other health problems, the proteins they encode and knowledge of their roles in amelogenesis, combining evidence from human phenotypes, inheritance patterns, mouse models, and in vitro studies. An LOVD resource (http://dna2.leeds.ac.uk/LOVD/) containing all published gene mutations for AI presenting in isolation of other health problems is described. We use this resource to identify trends in the genes and mutations reported to cause AI in the 270 families for which molecular diagnoses have been reported by 23rd May 2017. Finally we discuss the potential value of the translation of AI genetics to clinical care with improved patient pathways and speculate on the possibility of novel treatments and prevention strategies for AI.
Keywords: LOVD; Leiden Open Variant Database; ameloblasts; amelogenesis; amelogenesis genetics; amelogenesis imperfecta; biomineralization; enamel.
Figures
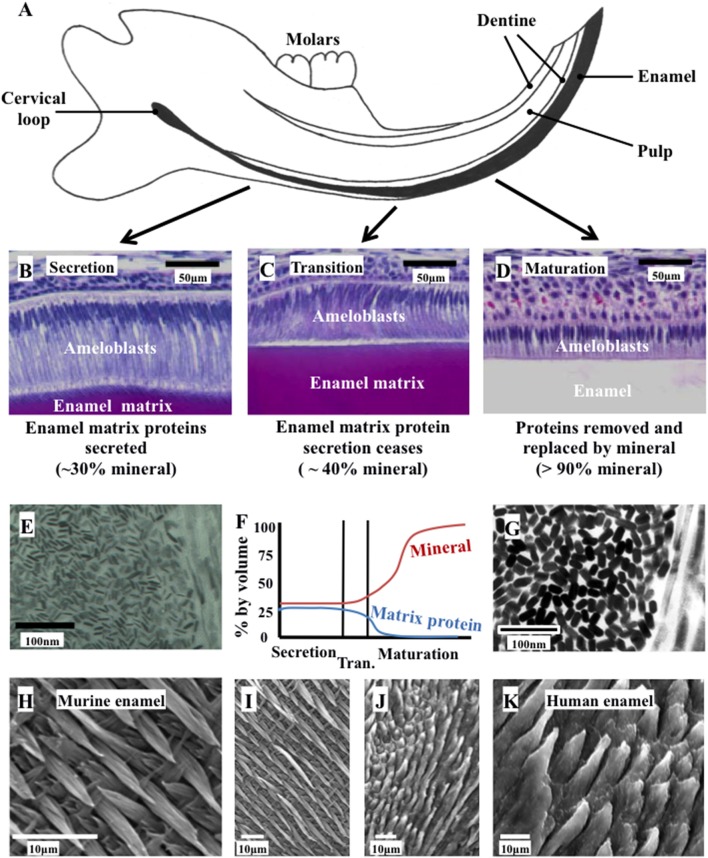
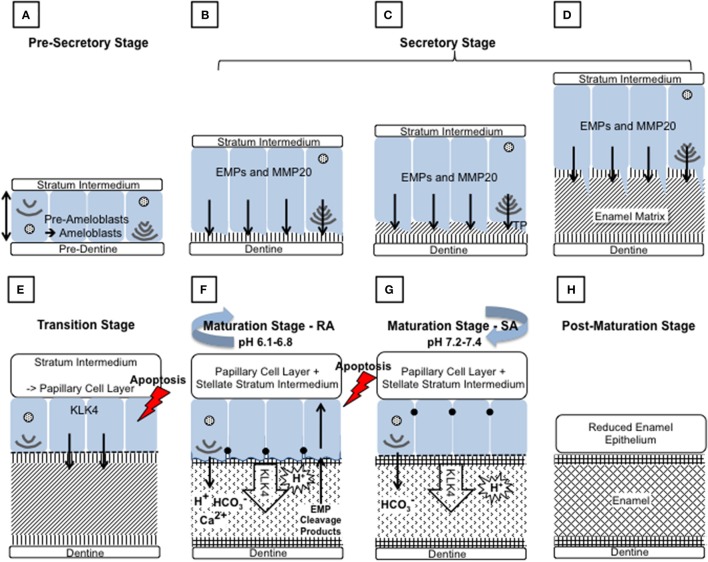
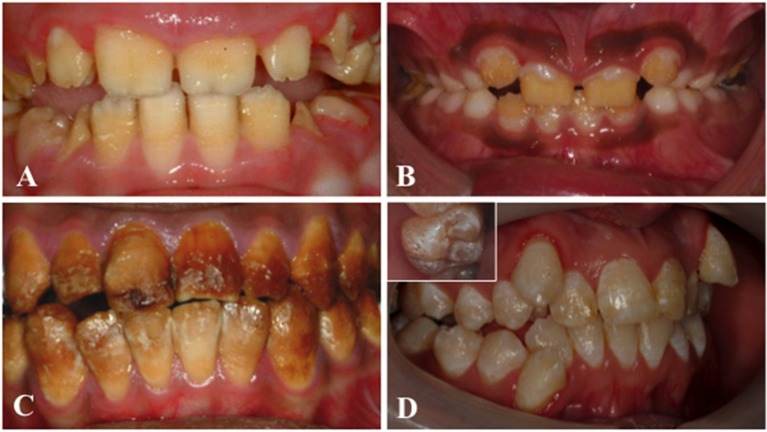
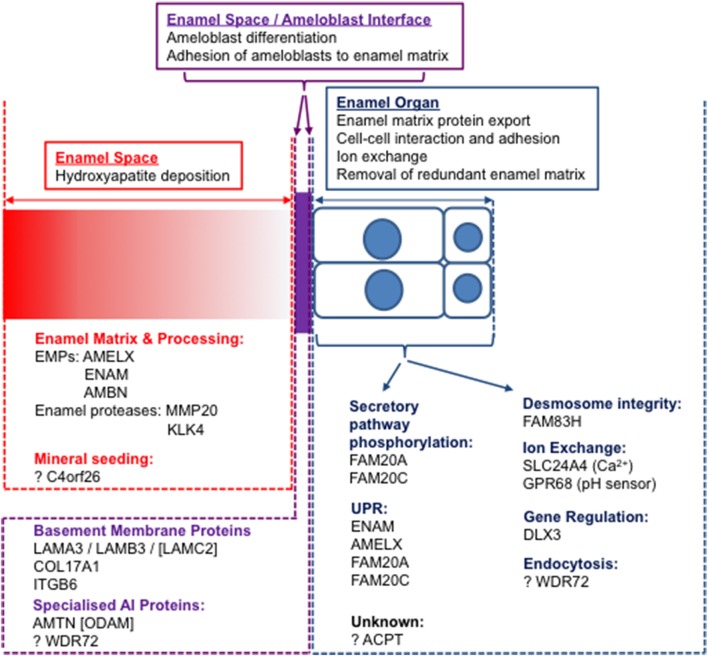
References
-
- Alberts B. J., Lewis J., Raff M., Roberts K., Walter P. (2002). Molecular Biology of the Cell. New York, NY: Garland Science.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources