

ADIPOQ/adiponectin induces cytotoxic autophagy in breast cancer cells through STK11/LKB1-mediated activation of the AMPK-ULK1 axis
- PMID: 28696138
- PMCID: PMC5584870
- DOI: 10.1080/15548627.2017.1332565
ADIPOQ/adiponectin induces cytotoxic autophagy in breast cancer cells through STK11/LKB1-mediated activation of the AMPK-ULK1 axis
Abstract
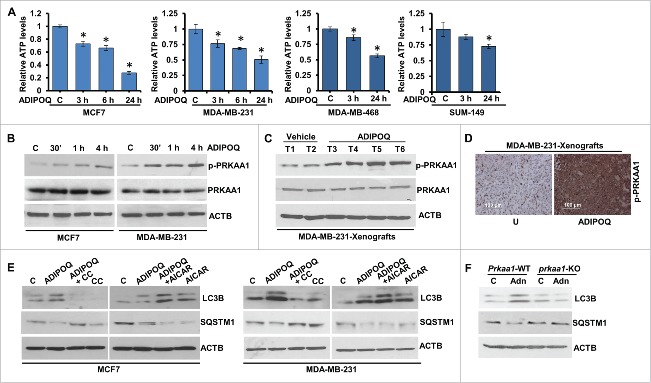
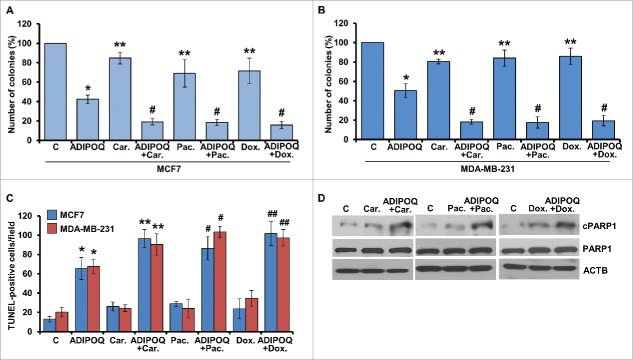
ADIPOQ/adiponectin, an adipocytokine secreted by adipocytes in the breast tumor microenvironment, negatively regulates cancer cell growth hence increased levels of ADIPOQ/adiponectin are associated with decreased breast cancer growth. However, its mechanisms of action remain largely elusive. We report that ADIPOQ/adiponectin induces a robust accumulation of autophagosomes, increases MAP1LC3B-II/LC3B-II and decreases SQSTM1/p62 in breast cancer cells. ADIPOQ/adiponectin-treated cells and xenografts exhibit increased expression of autophagy-related proteins. LysoTracker Red-staining and tandem-mCherry-GFP-LC3B assay show that fusion of autophagosomes and lysosomes is augmented upon ADIPOQ/adiponectin treatment. ADIPOQ/adiponectin significantly inhibits breast cancer growth and induces apoptosis both in vitro and in vivo, and these events are preceded by macroautophagy/autophagy, which is integral for ADIPOQ/adiponectin-mediated cell death. Accordingly, blunting autophagosome formation, blocking autophagosome-lysosome fusion or genetic-knockout of BECN1/Beclin1 and ATG7 effectively impedes ADIPOQ/adiponectin induced growth-inhibition and apoptosis-induction. Mechanistic studies show that ADIPOQ/adiponectin reduces intracellular ATP levels and increases PRKAA1 phosphorylation leading to ULK1 activation. AMPK-inhibition abrogates ADIPOQ/adiponectin-induced ULK1-activation, LC3B-turnover and SQSTM1/p62-degradation while AMPK-activation potentiates ADIPOQ/adiponectin's effects. Further, ADIPOQ/adiponectin-mediated AMPK-activation and autophagy-induction are regulated by upstream master-kinase STK11/LKB1, which is a key node in antitumor function of ADIPOQ/adiponectin as STK11/LKB1-knockout abrogates ADIPOQ/adiponectin-mediated inhibition of breast tumorigenesis and molecular analyses of tumors corroborate in vitro mechanistic findings. ADIPOQ/adiponectin increases the efficacy of chemotherapeutic agents. Notably, high expression of ADIPOQ receptor ADIPOR2, ADIPOQ/adiponectin and BECN1 significantly correlates with increased overall survival in chemotherapy-treated breast cancer patients. Collectively, these data uncover that ADIPOQ/adiponectin induces autophagic cell death in breast cancer and provide in vitro and in vivo evidence for the integral role of STK11/LKB1-AMPK-ULK1 axis in ADIPOQ/adiponectin-mediated cytotoxic autophagy.
Keywords: AMPK; STK11/LKB1; ULK1; adiponectin; autophagy; breast cancer.
Figures






References
-
- Vona-Davis L, Howard-McNatt M, Rose DP. Adiposity, type 2 diabetes and the metabolic syndrome in breast cancer. Obes Rev 2007; 8:395-408; PMID:17716297; https://doi.org/10.1111/j.1467-789X.2007.00396.x - DOI - PubMed
-
- Sharma D, Davidson NE. Obesity and breast cancer: a multipartite connection. J Mammary Gland Biol Neoplasia 2013; 18:253-5; PMID:24190309; https://doi.org/10.1007/s10911-013-9306-4 - DOI - PMC - PubMed
-
- Iyengar NM, Zhou XK, Gucalp A, Morris PG, Howe LR, Giri DD, Morrow M, Wang H, Pollak M, Jones LW, et al.. Systemic Correlates of White Adipose Tissue Inflammation in Early-Stage Breast Cancer. Clinical cancer research: an official journal of the American Association for Cancer Research 2016; 22:2283-89; PMID:26712688; https://doi.org/10.1158/1078-0432.CCR-15-2239 - DOI - PMC - PubMed
-
- Lee Y, Jung WH, Koo JS. Adipocytes can induce epithelial-mesenchymal transition in breast cancer cells. Breast cancer research and treatment 2015; 153:323-35; PMID:26285644; https://doi.org/10.1007/s10549-015-3550-9 - DOI - PubMed
-
- Santander AM, Lopez-Ocejo O, Casas O, Agostini T, Sanchez L, Lamas-Basulto E, Carrio R, Cleary MP, Gonzalez-Perez RR, Torroella-Kouri M. Paracrine Interactions between Adipocytes and Tumor Cells Recruit and Modify Macrophages to the Mammary Tumor Microenvironment: The Role of Obesity and Inflammation in Breast Adipose Tissue. Cancers 2015; 7:143-78; PMID:25599228; https://doi.org/10.3390/cancers7010143 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous