

A Lectin Receptor-Like Kinase Mediates Pattern-Triggered Salicylic Acid Signaling
- PMID: 28696275
- PMCID: PMC5543950
- DOI: 10.1104/pp.17.00404
A Lectin Receptor-Like Kinase Mediates Pattern-Triggered Salicylic Acid Signaling
Abstract
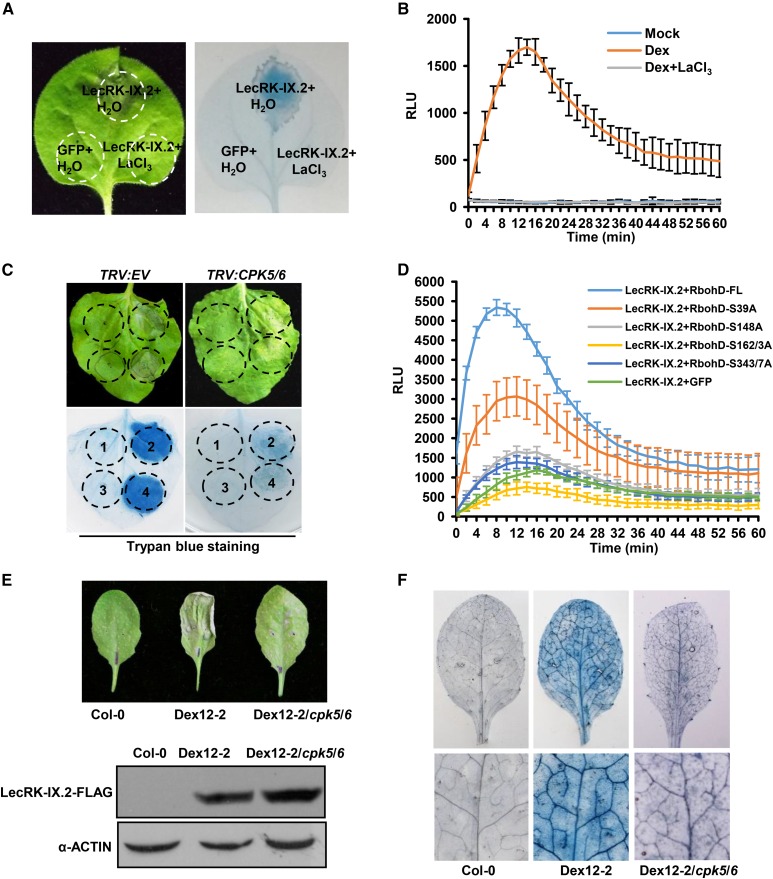
Plant surface-localized pathogen recognition receptors (PRRs) perceive conserved microbial features, termed pathogen-associated molecular patterns (PAMPs), resulting in disease resistance. PAMP perception leads to calcium influx, MAPK activation, a burst of reactive oxygen species (ROS) mediated by RbohD, accumulation of the defense hormone salicylic acid (SA), and callose deposition. Lectin receptor-like kinases (LecRKs) belong to a specific PRR family and are important players in plant innate immunity. Here, we report that LecRK-IX.2 is a positive regulator of PRR-triggered immunity. Pathogen infection activated the transcription of Arabidopsis (Arabidopsis thaliana) LecRK-IX.2, and the LecRK-IX.2 knockout lines exhibited enhanced susceptibility to virulent Pseudomonas syringae pv tomato DC3000. In addition, LecRK-IX.2 is capable of inducing RbohD phosphorylation, likely by recruiting calcium-dependent protein kinases to trigger ROS production in Arabidopsis. Overexpression of LecRK-IX.2 resulted in elevated ROS and SA and enhanced systemic acquired resistance to P. syringae pv tomato DC3000. Our data highlight the importance of LecRKs in plant immune signaling and SA accumulation.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures








Similar articles
-
The Arabidopsis lectin receptor kinase LecRK-V.5 represses stomatal immunity induced by Pseudomonas syringae pv. tomato DC3000.PLoS Pathog. 2012 Feb;8(2):e1002513. doi: 10.1371/journal.ppat.1002513. Epub 2012 Feb 9. PLoS Pathog. 2012. PMID: 22346749 Free PMC article.
-
The Arabidopsis thaliana lectin receptor kinase LecRK-I.9 is required for full resistance to Pseudomonas syringae and affects jasmonate signalling.Mol Plant Pathol. 2017 Sep;18(7):937-948. doi: 10.1111/mpp.12457. Epub 2016 Sep 15. Mol Plant Pathol. 2017. PMID: 27399963 Free PMC article.
-
The receptor-like cytoplasmic kinase PCRK1 contributes to pattern-triggered immunity against Pseudomonas syringae in Arabidopsis thaliana.New Phytol. 2015 Jul;207(1):78-90. doi: 10.1111/nph.13345. Epub 2015 Feb 25. New Phytol. 2015. PMID: 25711411
-
Regulation of the NADPH Oxidase RBOHD During Plant Immunity.Plant Cell Physiol. 2015 Aug;56(8):1472-80. doi: 10.1093/pcp/pcv063. Epub 2015 May 4. Plant Cell Physiol. 2015. PMID: 25941234 Review.
-
Chitin receptor-mediated activation of MAP kinases and ROS production in rice and Arabidopsis.Plant Signal Behav. 2017 Sep 2;12(9):e1361076. doi: 10.1080/15592324.2017.1361076. Epub 2017 Aug 14. Plant Signal Behav. 2017. PMID: 28805500 Free PMC article. Review.
Cited by
-
Combined transcriptome and metabolome analysis identifies defence responses in spider mite-infested pepper (Capsicum annuum).J Exp Bot. 2020 Jan 1;71(1):330-343. doi: 10.1093/jxb/erz422. J Exp Bot. 2020. PMID: 31557301 Free PMC article.
-
Computational prediction and in vitro analysis of the potential ligand binding site within the extracellular ATP receptor, P2K2.Plant Signal Behav. 2023 Dec 31;18(1):2173146. doi: 10.1080/15592324.2023.2173146. Plant Signal Behav. 2023. PMID: 36723515 Free PMC article.
-
The Arabidopsis thaliana onset of leaf death 12 mutation in the lectin receptor kinase P2K2 results in an autoimmune phenotype.BMC Plant Biol. 2023 Jun 2;23(1):294. doi: 10.1186/s12870-023-04300-0. BMC Plant Biol. 2023. PMID: 37264342 Free PMC article.
-
Multi-stress resilience in plants recovering from submergence.Plant Biotechnol J. 2023 Mar;21(3):466-481. doi: 10.1111/pbi.13944. Epub 2022 Nov 14. Plant Biotechnol J. 2023. PMID: 36217562 Free PMC article. Review.
-
Ligand recognition and signal transduction by lectin receptor-like kinases in plant immunity.Front Plant Sci. 2023 Jun 15;14:1201805. doi: 10.3389/fpls.2023.1201805. eCollection 2023. Front Plant Sci. 2023. PMID: 37396638 Free PMC article. Review.
References
-
- Balagué C, Gouget A, Bouchez O, Souriac C, Haget N, Boutet-Mercey S, Govers F, Roby D, Canut H (2016) The Arabidopsis thaliana lectin receptor kinase LecRK-I.9 is required for full resistance to Pseudomonas syringae and affects jasmonate signalling. Mol Plant Pathol doi/10.1111/mpp.12457 - DOI - PMC - PubMed
-
- Bent A. (2006) Arabidopsis thaliana floral dip transformation method. Methods Mol Biol 343: 87–103 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
