

Escherichia coli cytochrome c peroxidase is a respiratory oxidase that enables the use of hydrogen peroxide as a terminal electron acceptor
- PMID: 28696311
- PMCID: PMC5565418
- DOI: 10.1073/pnas.1701587114
Escherichia coli cytochrome c peroxidase is a respiratory oxidase that enables the use of hydrogen peroxide as a terminal electron acceptor
Abstract
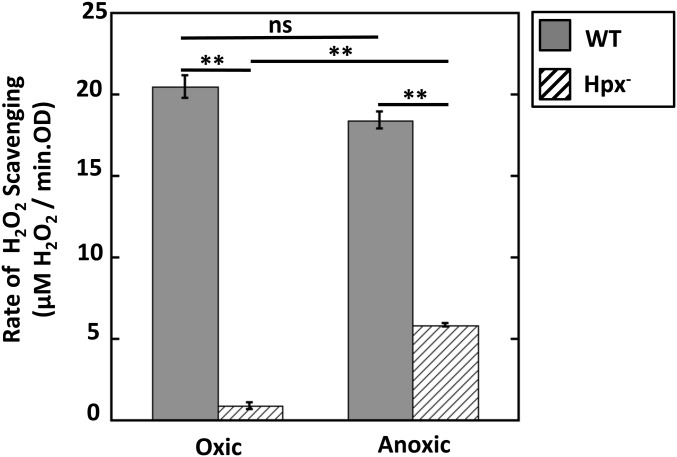
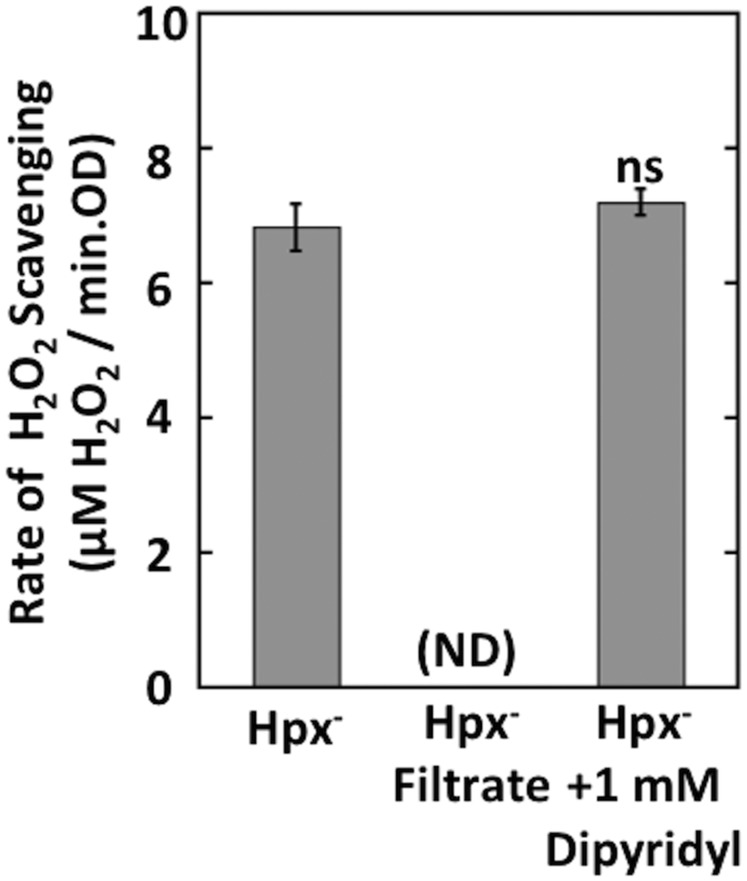
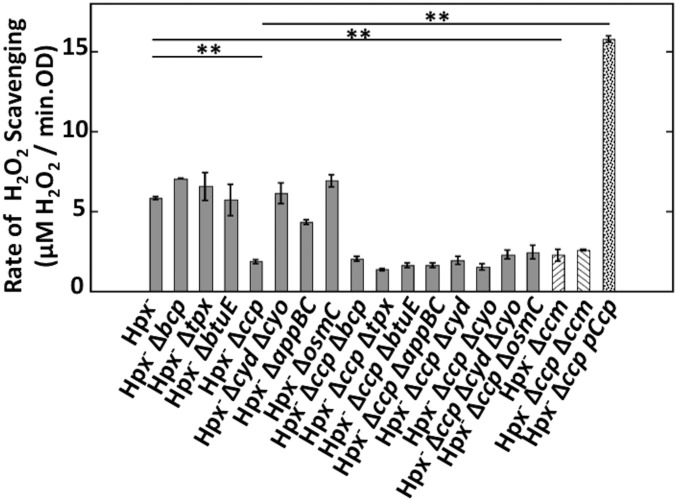
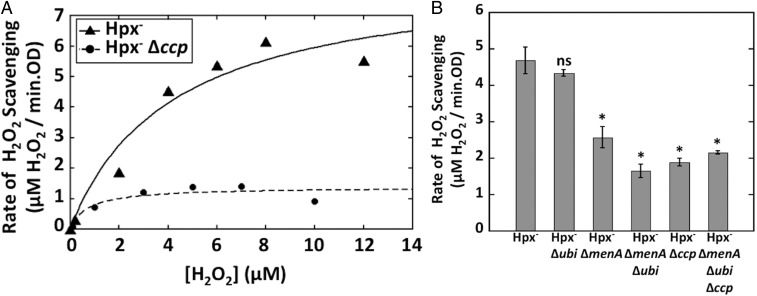
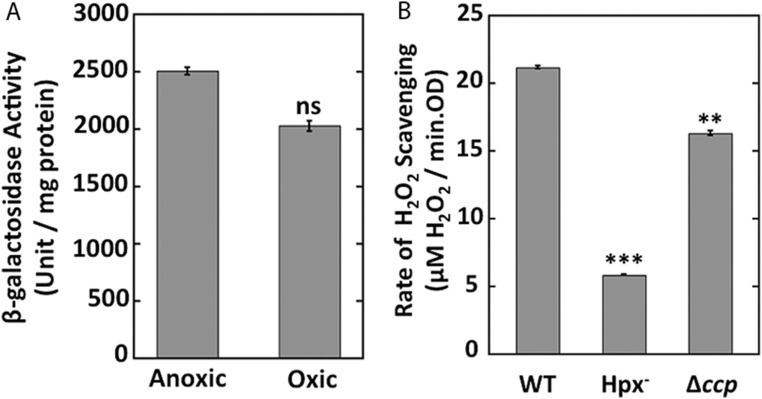
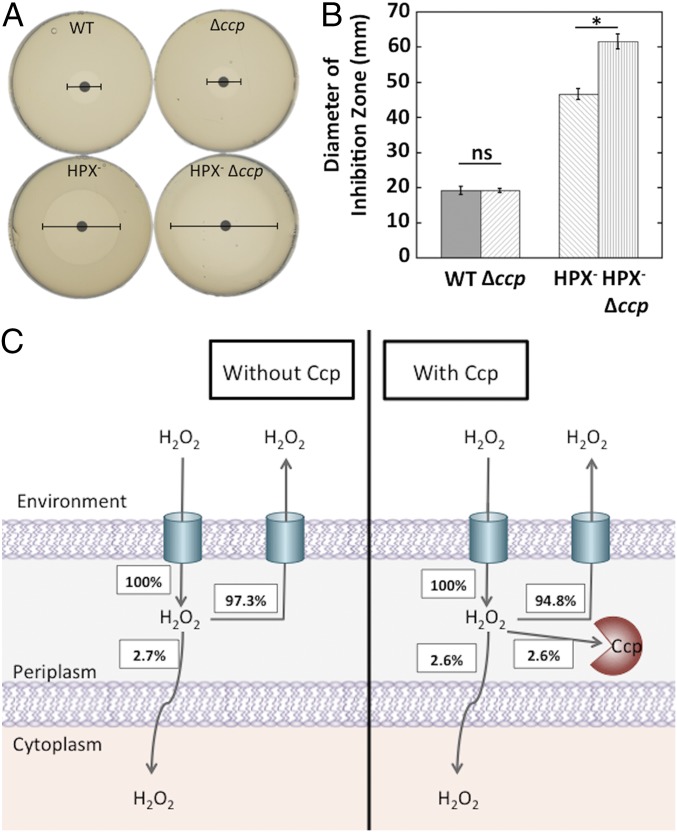
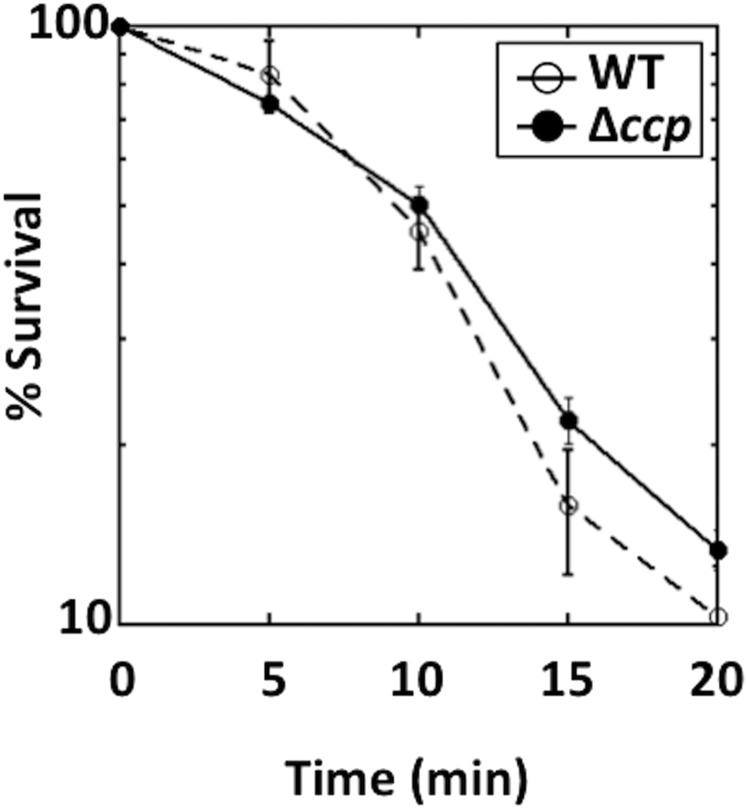
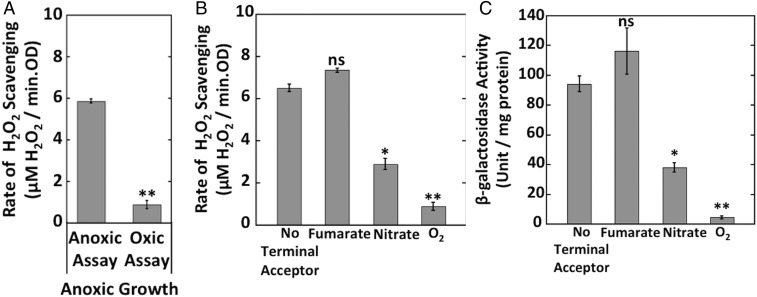
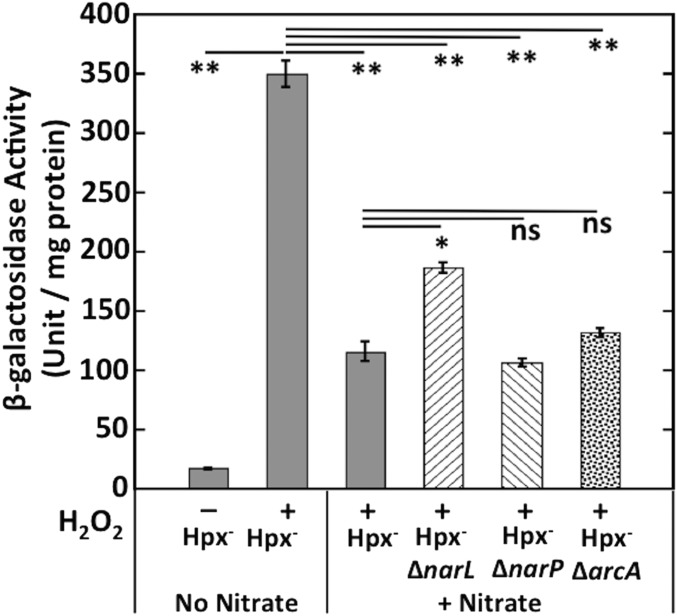
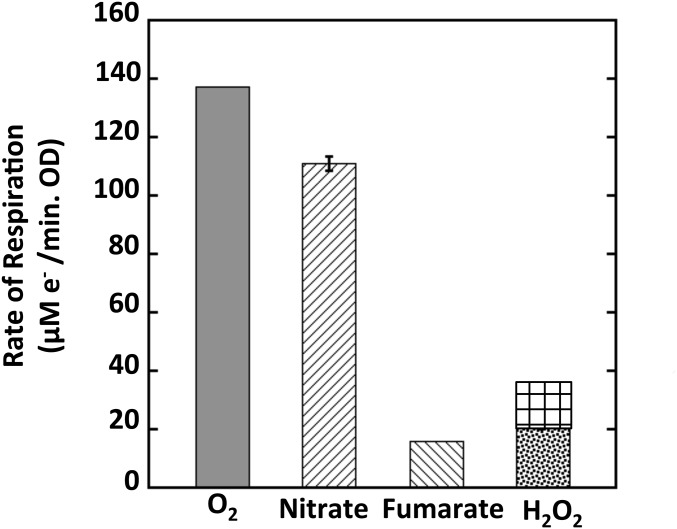
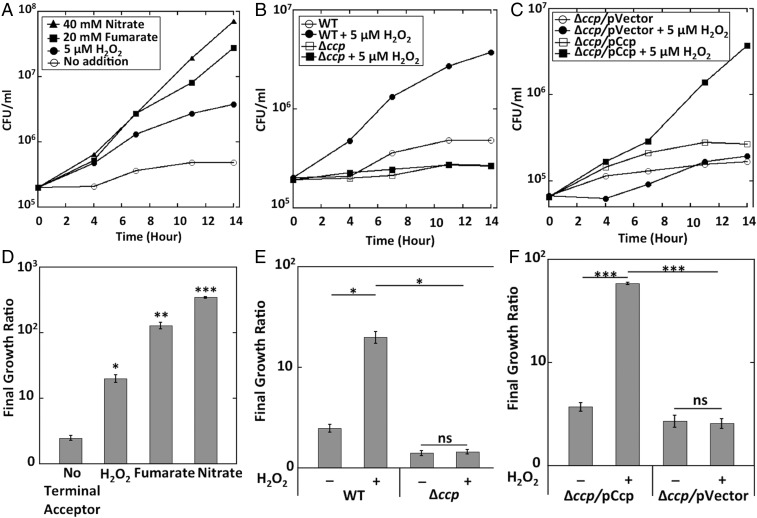
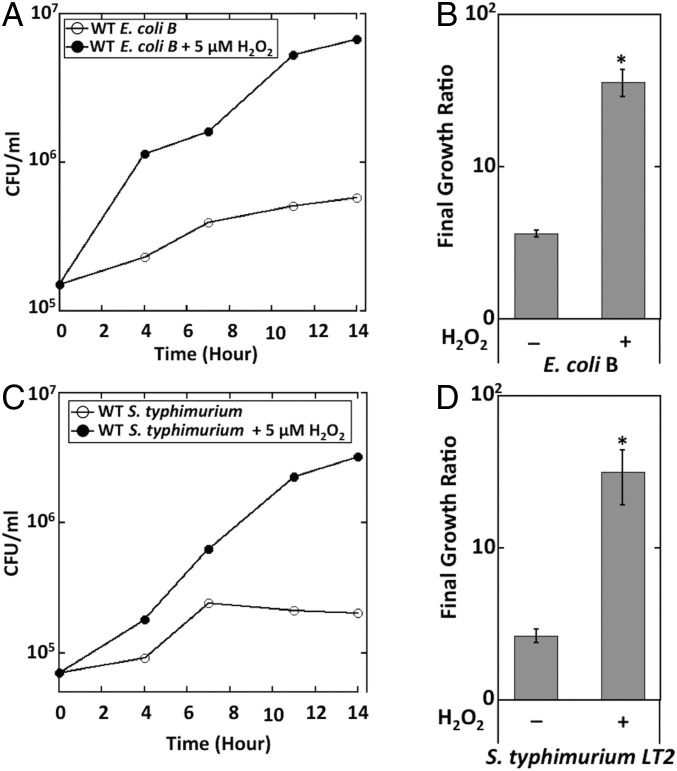
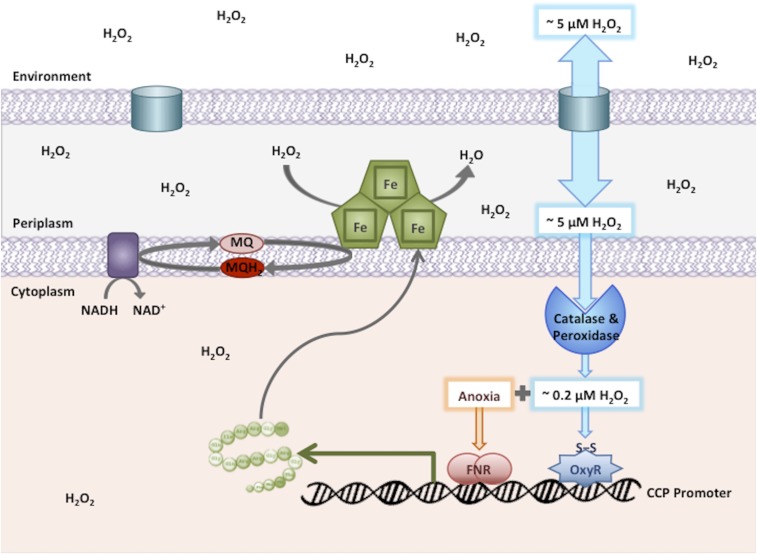
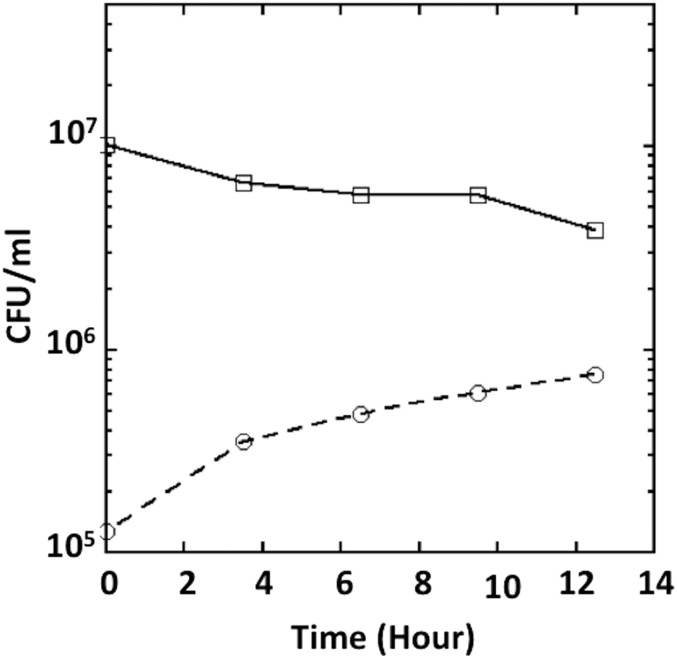
Microbial cytochrome c peroxidases (Ccp) have been studied for 75 years, but their physiological roles are unclear. Ccps are located in the periplasms of bacteria and the mitochondrial intermembrane spaces of fungi. In this study, Ccp is demonstrated to be a significant degrader of hydrogen peroxide in anoxic Escherichia coli Intriguingly, ccp transcription requires both the presence of H2O2 and the absence of O2 Experiments show that Ccp lacks enough activity to shield the cytoplasm from exogenous H2O2 However, it receives electrons from the quinone pool, and its flux rate approximates flow to other anaerobic electron acceptors. Indeed, Ccp enabled E. coli to grow on a nonfermentable carbon source when H2O2 was supplied. Salmonella behaved similarly. This role rationalizes ccp repression in oxic environments. We speculate that micromolar H2O2 is created both biologically and abiotically at natural oxic/anoxic interfaces. The OxyR response appears to exploit this H2O2 as a terminal oxidant while simultaneously defending the cell against its toxicity.
Keywords: OxyR; anaerobic respiration; oxidative stress.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Cytochrome c peroxidase facilitates the beneficial use of H2O2 in prokaryotes.Proc Natl Acad Sci U S A. 2017 Aug 15;114(33):8678-8680. doi: 10.1073/pnas.1710943114. Epub 2017 Aug 1. Proc Natl Acad Sci U S A. 2017. PMID: 28765365 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
