

A Tubulin Binding Switch Underlies Kip3/Kinesin-8 Depolymerase Activity
- PMID: 28697331
- PMCID: PMC5573156
- DOI: 10.1016/j.devcel.2017.06.011
A Tubulin Binding Switch Underlies Kip3/Kinesin-8 Depolymerase Activity
Abstract
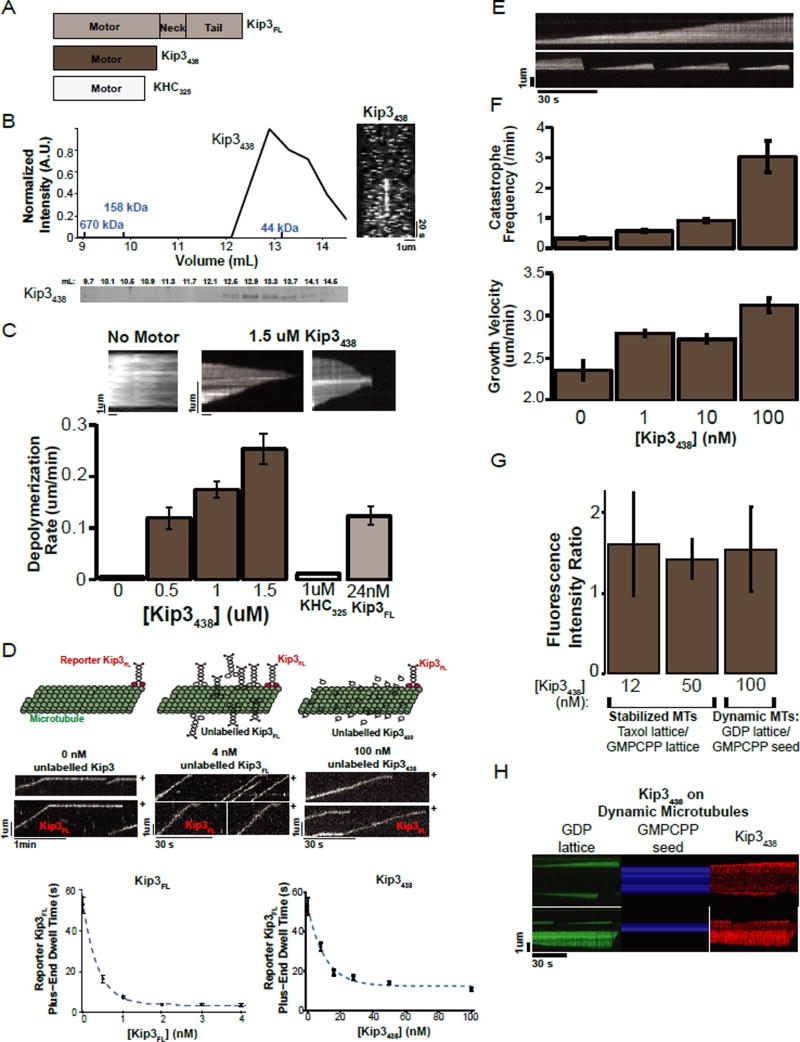
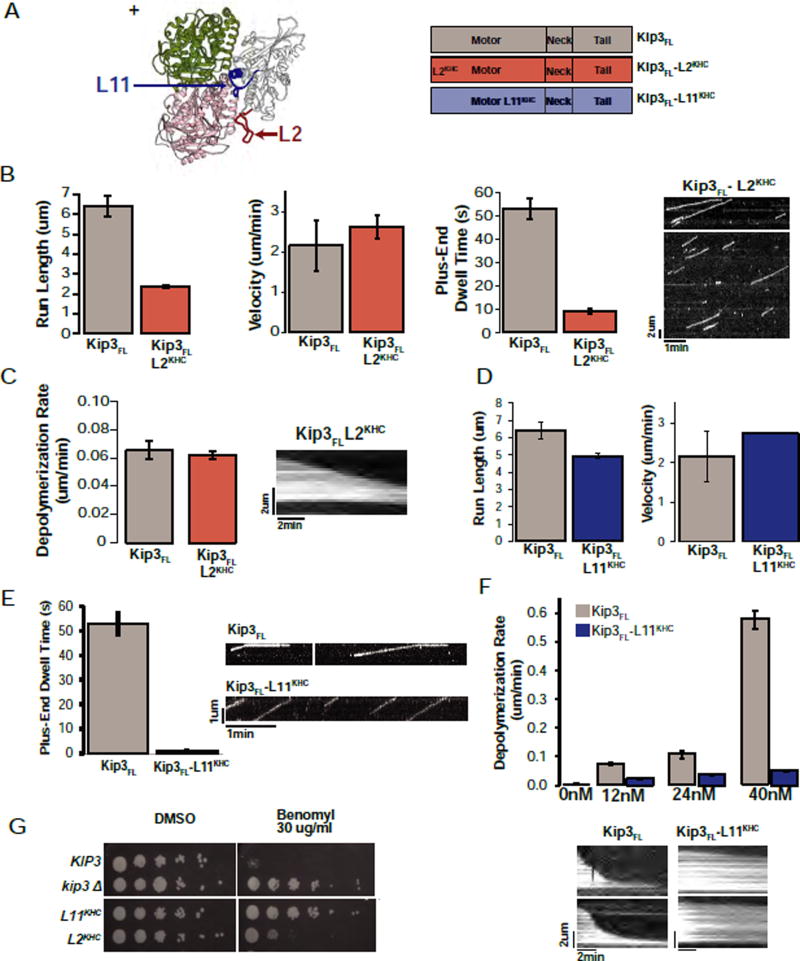
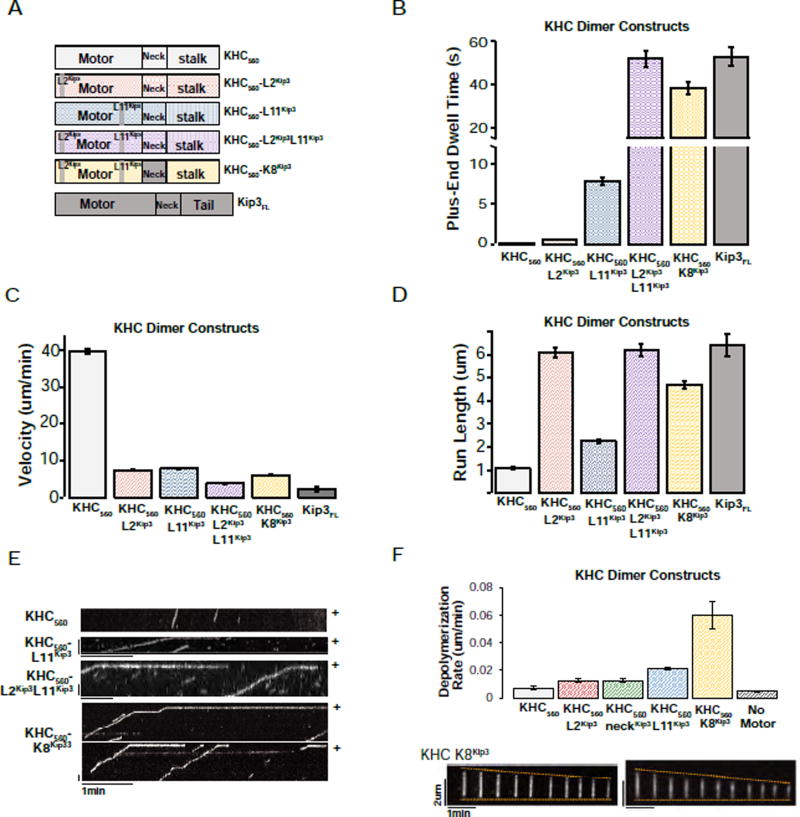
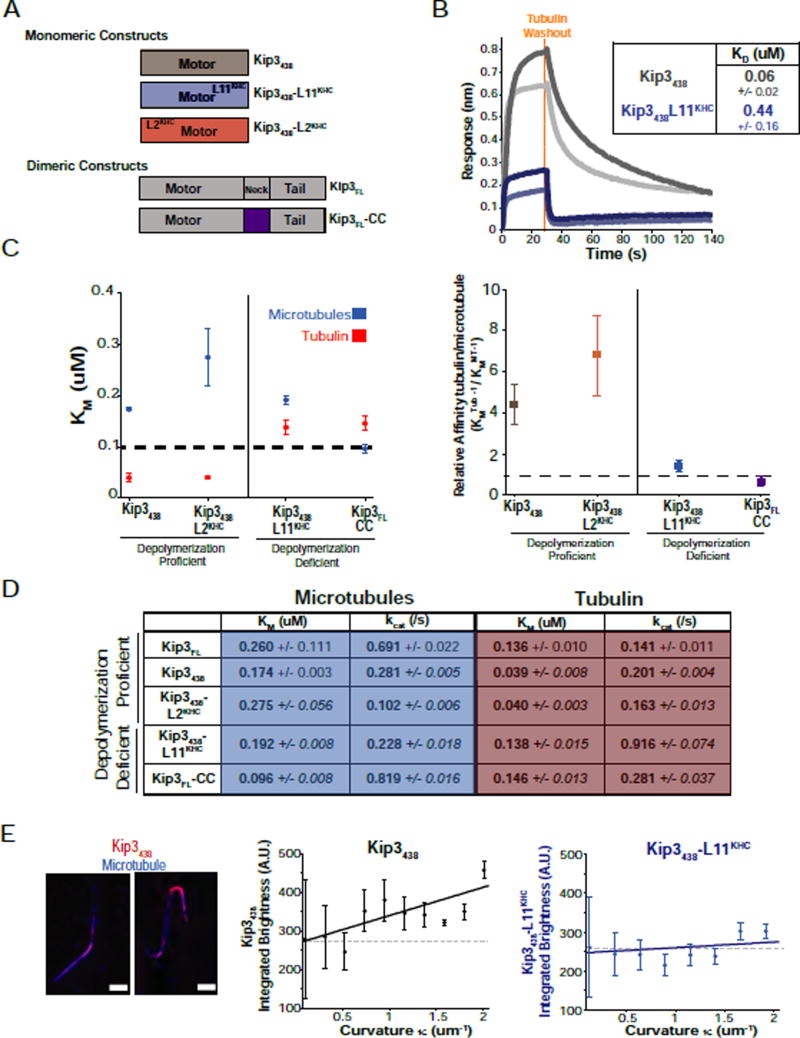
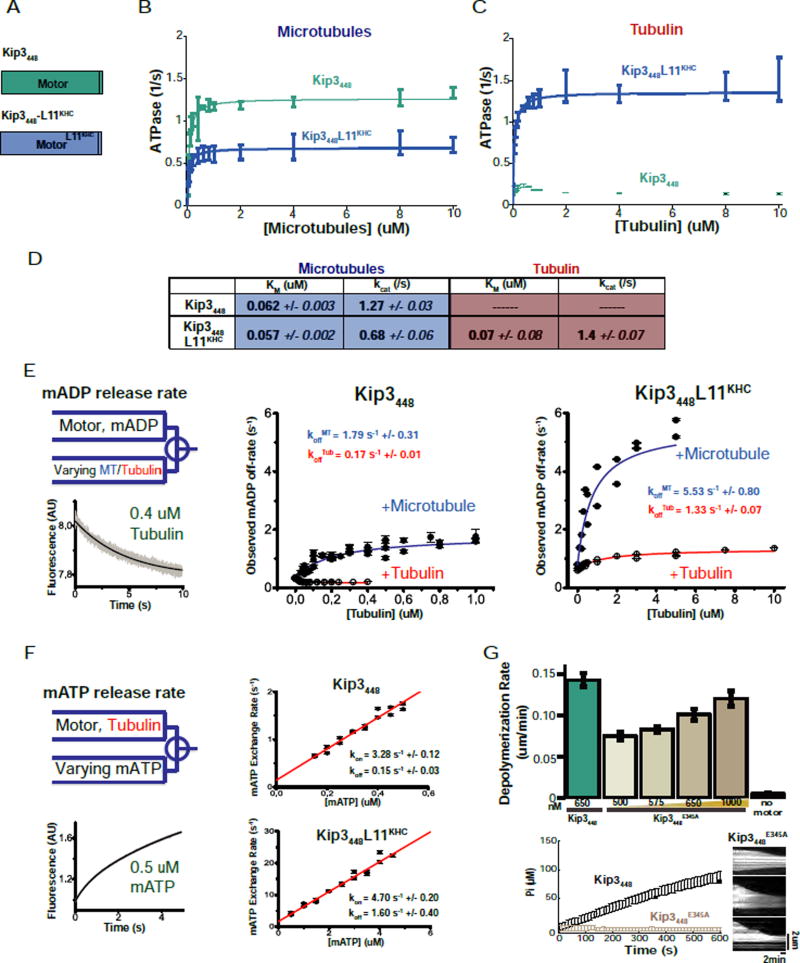
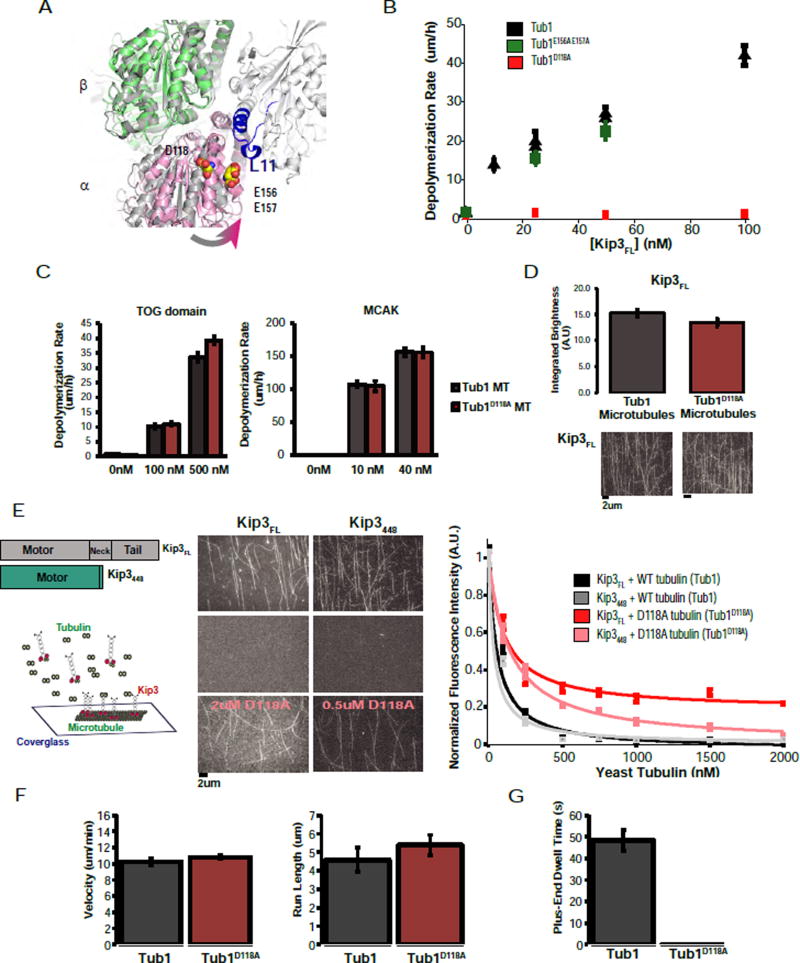
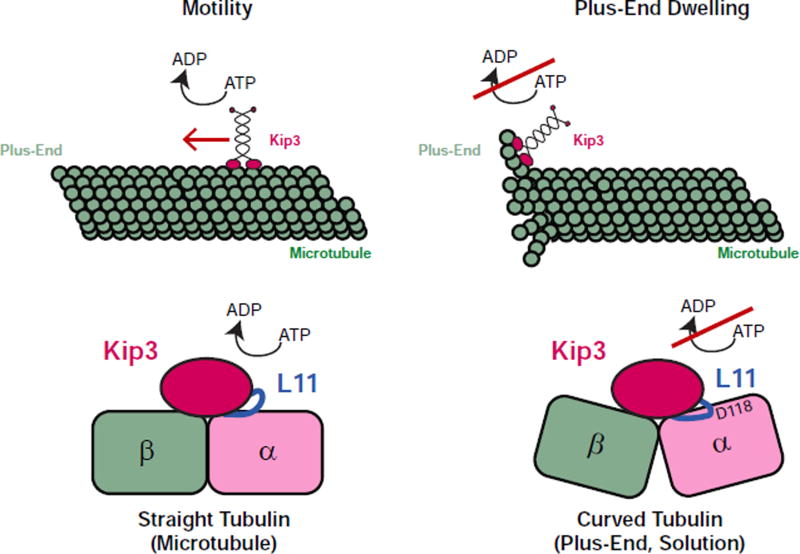
Kinesin-8 motors regulate the size of microtubule structures, using length-dependent accumulation at the plus end to preferentially disassemble long microtubules. Despite extensive study, the kinesin-8 depolymerase mechanism remains under debate. Here, we provide evidence for an alternative, tubulin curvature-sensing model of microtubule depolymerization by the budding yeast kinesin-8, Kip3. Kinesin-8/Kip3 uses ATP hydrolysis, like other kinesins, for stepping on the microtubule lattice, but at the plus end Kip3 undergoes a switch: its ATPase activity is suppressed when it binds tightly to the curved conformation of tubulin. This prolongs plus-end binding, stabilizes protofilament curvature, and ultimately promotes microtubule disassembly. The tubulin curvature-sensing model is supported by our identification of Kip3 structural elements necessary and sufficient for plus-end binding and depolymerase activity, as well as by the identification of an α-tubulin residue specifically required for the Kip3-curved tubulin interaction. Together, these findings elucidate a major regulatory mechanism controlling the size of cellular microtubule structures.
Keywords: depolymerization; kinesins; microtubule associated proteins; microtubule dynamics; spindle scaling.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Bechstedt S, Lu K, Brouhard Gary J. Doublecortin Recognizes the Longitudinal Curvature of the Microtubule End and Lattice. Current Biology. 2014;24:2366–2375. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
