

DGAT1-Dependent Lipid Droplet Biogenesis Protects Mitochondrial Function during Starvation-Induced Autophagy
- PMID: 28697336
- PMCID: PMC5553613
- DOI: 10.1016/j.devcel.2017.06.003
DGAT1-Dependent Lipid Droplet Biogenesis Protects Mitochondrial Function during Starvation-Induced Autophagy
Abstract
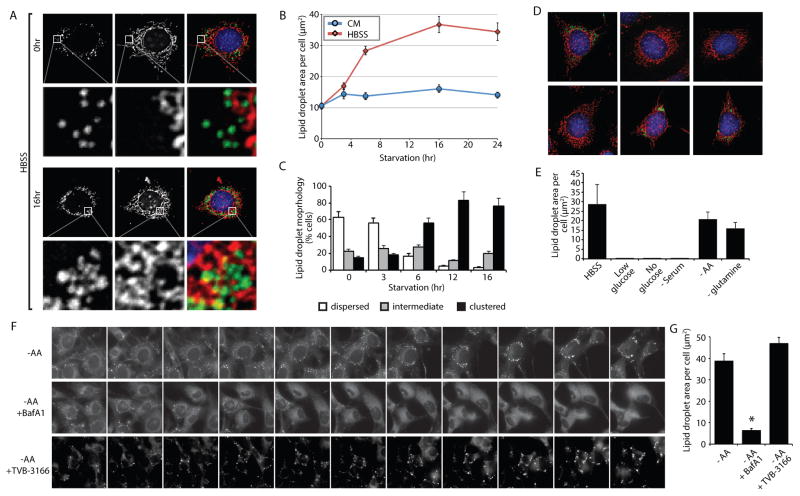
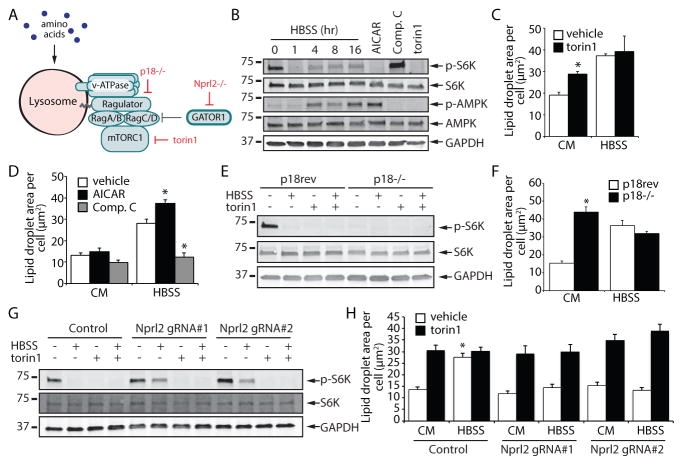
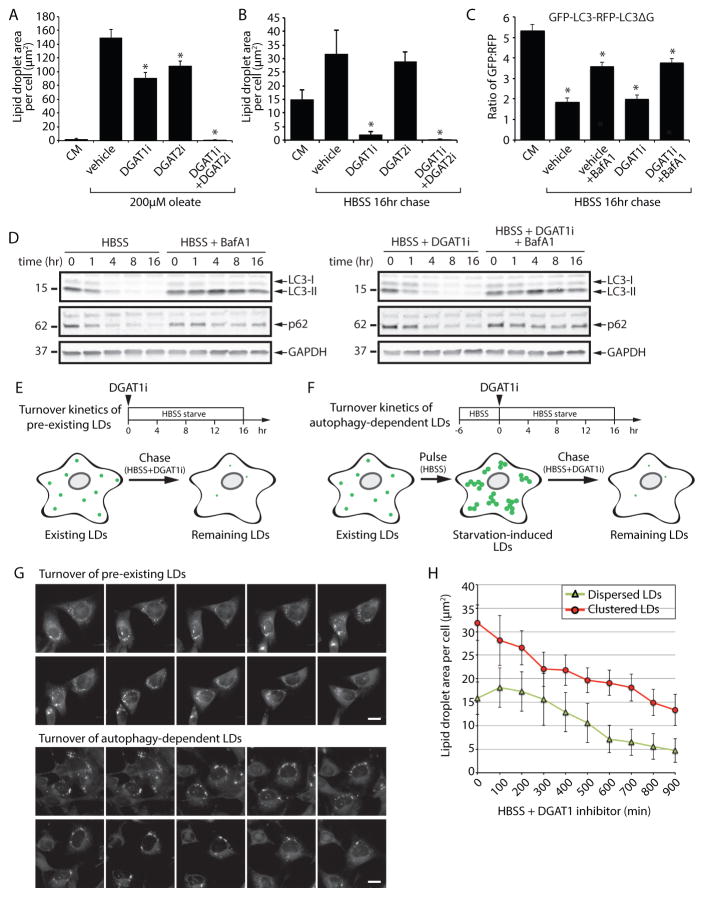
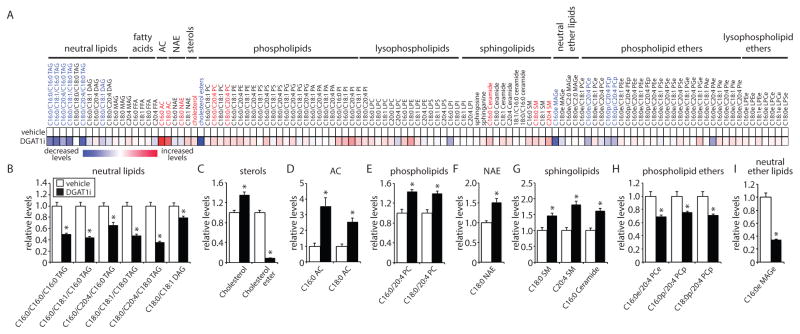
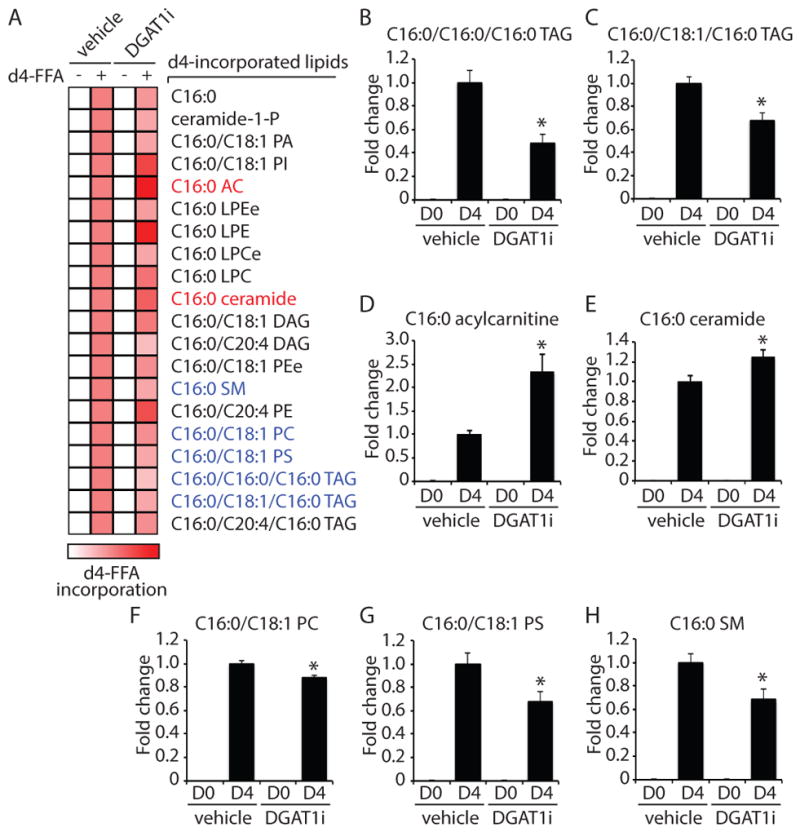
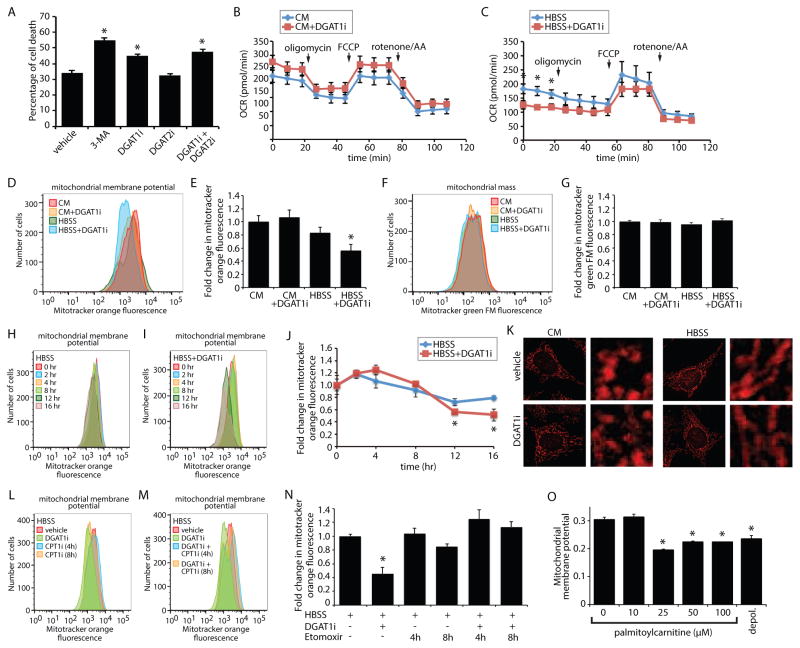
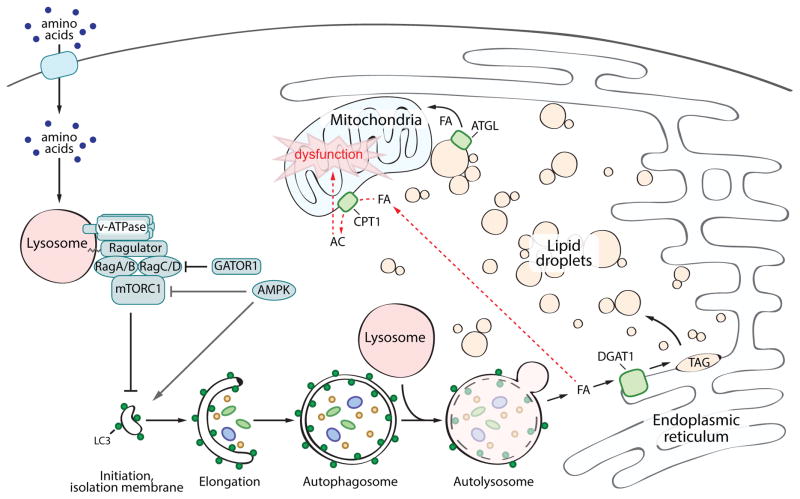
Lipid droplets (LDs) provide an "on-demand" source of fatty acids (FAs) that can be mobilized in response to fluctuations in nutrient abundance. Surprisingly, the amount of LDs increases during prolonged periods of nutrient deprivation. Why cells store FAs in LDs during an energy crisis is unknown. Our data demonstrate that mTORC1-regulated autophagy is necessary and sufficient for starvation-induced LD biogenesis. The ER-resident diacylglycerol acyltransferase 1 (DGAT1) selectively channels autophagy-liberated FAs into new, clustered LDs that are in close proximity to mitochondria and are lipolytically degraded. However, LDs are not required for FA delivery to mitochondria but instead function to prevent acylcarnitine accumulation and lipotoxic dysregulation of mitochondria. Our data support a model in which LDs provide a lipid buffering system that sequesters FAs released during the autophagic degradation of membranous organelles, reducing lipotoxicity. These findings reveal an unrecognized aspect of the cellular adaptive response to starvation, mediated by LDs.
Keywords: ATGL; DGAT1; DGAT2; autophagy; lipid droplet; lipotoxicity; mTORC1; mitochondria; starvation; triacylglycerol.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Lipid Droplets Guard Mitochondria during Autophagy.Dev Cell. 2017 Jul 10;42(1):1-2. doi: 10.1016/j.devcel.2017.06.018. Dev Cell. 2017. PMID: 28697329
-
Recycling the danger via lipid droplet biogenesis after autophagy.Autophagy. 2017;13(11):1995-1997. doi: 10.1080/15548627.2017.1371394. Epub 2017 Oct 4. Autophagy. 2017. PMID: 28873005 Free PMC article.
References
-
- Beavis AD, Brannan RD, Garlid KD. Swelling and contraction of the mitochondrial matrix. I. A structural interpretation of the relationship between light scattering and matrix volume. J Biol Chem. 1985;260:13424–13433. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
