

A Novel Dual Eigen-Analysis of Mouse Multi-Tissues' Expression Profiles Unveils New Perspectives into Type 2 Diabetes
- PMID: 28698587
- PMCID: PMC5506042
- DOI: 10.1038/s41598-017-05405-x
A Novel Dual Eigen-Analysis of Mouse Multi-Tissues' Expression Profiles Unveils New Perspectives into Type 2 Diabetes
Abstract
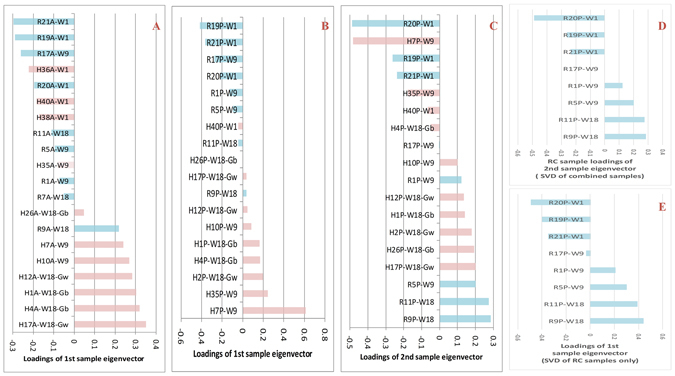
Type 2 diabetes (T2D) is a complex and polygenic disease yet in need of a complete picture of its development mechanisms. To better understand the mechanisms, we examined gene expression profiles of multi-tissues from outbred mice fed with a high-fat diet (HFD) or regular chow at weeks 1, 9, and 18. To analyze such complex data, we proposed a novel dual eigen-analysis, in which the sample- and gene-eigenvectors correspond respectively to the macro- and micro-biology information. The dual eigen-analysis identified the HFD eigenvectors as well as the endogenous eigenvectors for each tissue. The results imply that HFD influences the hepatic function or the pancreatic development as an exogenous factor, while in adipose HFD's impact roughly coincides with the endogenous eigenvector driven by aging. The enrichment analysis of the eigenvectors revealed diverse HFD impact on the three tissues over time. The diversity includes: inflammation, degradation of branched chain amino acids (BCAA), and regulation of peroxisome proliferator activated receptor gamma (PPARγ). We reported that in the pancreas remarkable up-regulation of angiogenesis as downstream of the HIF signaling pathway precedes hyperinsulinemia. The dual eigen-analysis and discoveries provide new evaluations/guidance in T2D prevention and therapy, and will also promote new thinking in biology and medicine.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures





Similar articles
-
Adiponectin is expressed in the pancreas of high-fat-diet-fed mice and protects pancreatic endothelial function during the development of type 2 diabetes.Diabetes Metab. 2014 Nov;40(5):363-72. doi: 10.1016/j.diabet.2014.05.004. Epub 2014 Jun 27. Diabetes Metab. 2014. PMID: 24986510
-
Effects of a Lactobacillus paracasei B21060 based synbiotic on steatosis, insulin signaling and toll-like receptor expression in rats fed a high-fat diet.J Nutr Biochem. 2014 Jan;25(1):81-90. doi: 10.1016/j.jnutbio.2013.09.006. Epub 2013 Oct 10. J Nutr Biochem. 2014. PMID: 24314869
-
Antidiabetic effects of rice hull smoke extract on glucose-regulating mechanism in type 2 diabetic mice.J Agric Food Chem. 2012 Aug 1;60(30):7442-9. doi: 10.1021/jf3017749. Epub 2012 Jul 17. J Agric Food Chem. 2012. PMID: 22803686
-
Impaired adiponectin signaling contributes to disturbed catabolism of branched-chain amino acids in diabetic mice.Diabetes. 2015 Jan;64(1):49-59. doi: 10.2337/db14-0312. Epub 2014 Jul 28. Diabetes. 2015. PMID: 25071024
-
Diabetes and branched-chain amino acids: What is the link?J Diabetes. 2018 May;10(5):350-352. doi: 10.1111/1753-0407.12645. Epub 2018 Feb 13. J Diabetes. 2018. PMID: 29369529
Cited by
-
Identification of lysosomal lipolysis as an essential noncanonical mediator of adipocyte fasting and cold-induced lipolysis.J Clin Invest. 2025 Mar 17;135(6):e185340. doi: 10.1172/JCI185340. J Clin Invest. 2025. PMID: 40091840 Free PMC article.
-
A four eigen-phase model of multi-omics unveils new insights into yeast metabolic cycle.NAR Genom Bioinform. 2025 Mar 19;7(1):lqaf022. doi: 10.1093/nargab/lqaf022. eCollection 2025 Mar. NAR Genom Bioinform. 2025. PMID: 40109351 Free PMC article.
-
A data integration approach unveils a transcriptional signature of type 2 diabetes progression in rat and human islets.PLoS One. 2023 Oct 10;18(10):e0292579. doi: 10.1371/journal.pone.0292579. eCollection 2023. PLoS One. 2023. PMID: 37816033 Free PMC article.
-
TransBic: bucket trend-preserving biclustering for finding local and interpretable expression patterns.Brief Bioinform. 2024 Nov 22;26(1):bbaf050. doi: 10.1093/bib/bbaf050. Brief Bioinform. 2024. PMID: 39905952 Free PMC article.
-
MUREN: a robust and multi-reference approach of RNA-seq transcript normalization.BMC Bioinformatics. 2021 Jul 28;22(1):386. doi: 10.1186/s12859-021-04288-0. BMC Bioinformatics. 2021. PMID: 34320923 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
