

BCAT1 controls metabolic reprogramming in activated human macrophages and is associated with inflammatory diseases
- PMID: 28699638
- PMCID: PMC5510229
- DOI: 10.1038/ncomms16040
BCAT1 controls metabolic reprogramming in activated human macrophages and is associated with inflammatory diseases
Abstract
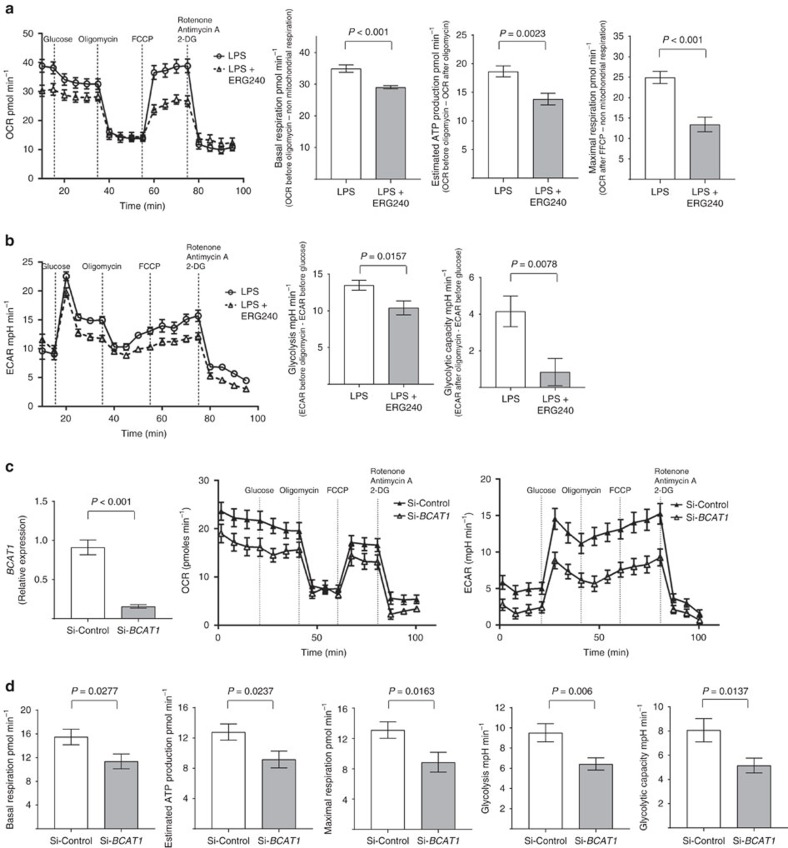
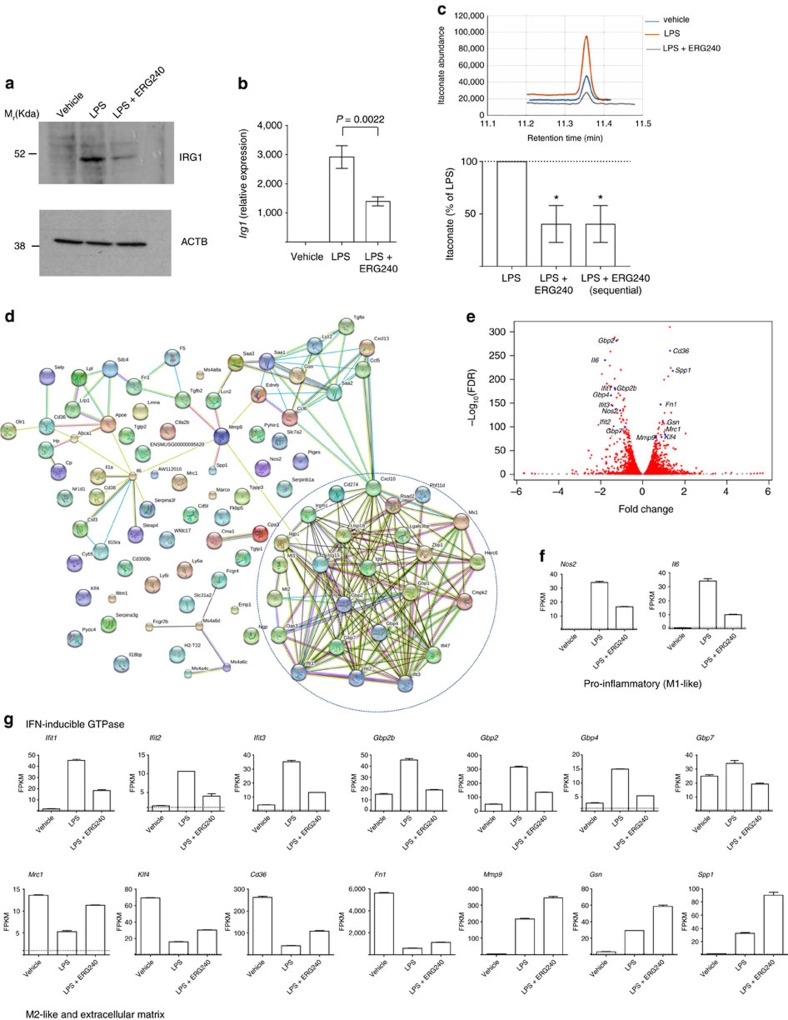
Branched-chain aminotransferases (BCAT) are enzymes that initiate the catabolism of branched-chain amino acids (BCAA), such as leucine, thereby providing macromolecule precursors; however, the function of BCATs in macrophages is unknown. Here we show that BCAT1 is the predominant BCAT isoform in human primary macrophages. We identify ERG240 as a leucine analogue that blocks BCAT1 activity. Selective inhibition of BCAT1 activity results in decreased oxygen consumption and glycolysis. This decrease is associated with reduced IRG1 levels and itaconate synthesis, suggesting involvement of BCAA catabolism through the IRG1/itaconate axis within the tricarboxylic acid cycle in activated macrophages. ERG240 suppresses production of IRG1 and itaconate in mice and contributes to a less proinflammatory transcriptome signature. Oral administration of ERG240 reduces the severity of collagen-induced arthritis in mice and crescentic glomerulonephritis in rats, in part by decreasing macrophage infiltration. These results establish a regulatory role for BCAT1 in macrophage function with therapeutic implications for inflammatory conditions.
Conflict of interest statement
A.E.P. is an employee of Ergon Pharmaceuticals Ltd. The remaining authors declare no competing financial interests.
Figures




References
-
- Brosnan J. T. & Brosnan M. E. Branched-chain amino acids: enzyme and substrate regulation. J. Nutr. 136, 207S–211S (2006). - PubMed
-
- Tom A. & Nair K. S. Assessment of branched-chain amino Acid status and potential for biomarkers. J. Nutr. 136, 324S–330S (2006). - PubMed
-
- Hutson S. M., Lieth E. & LaNoue K. F. Function of leucine in excitatory neurotransmitter metabolism in the central nervous system. J. Nutr. 131, 846S–850S (2001). - PubMed
-
- Sweatt A. J. et al.. Branched-chain amino acid catabolism: unique segregation of pathway enzymes in organ systems and peripheral nerves. Am. J. Physiol. Endocrinol. Metab. 286, E64–E76 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
