

A novel mechanism for mechanosensory-based rheotaxis in larval zebrafish
- PMID: 28700578
- PMCID: PMC5873946
- DOI: 10.1038/nature23014
A novel mechanism for mechanosensory-based rheotaxis in larval zebrafish
Erratum in
-
Erratum: A novel mechanism for mechanosensory-based rheotaxis in larval zebrafish.Nature. 2017 Sep 13;549(7671):292. doi: 10.1038/nature23895. Nature. 2017. PMID: 28905914
Abstract
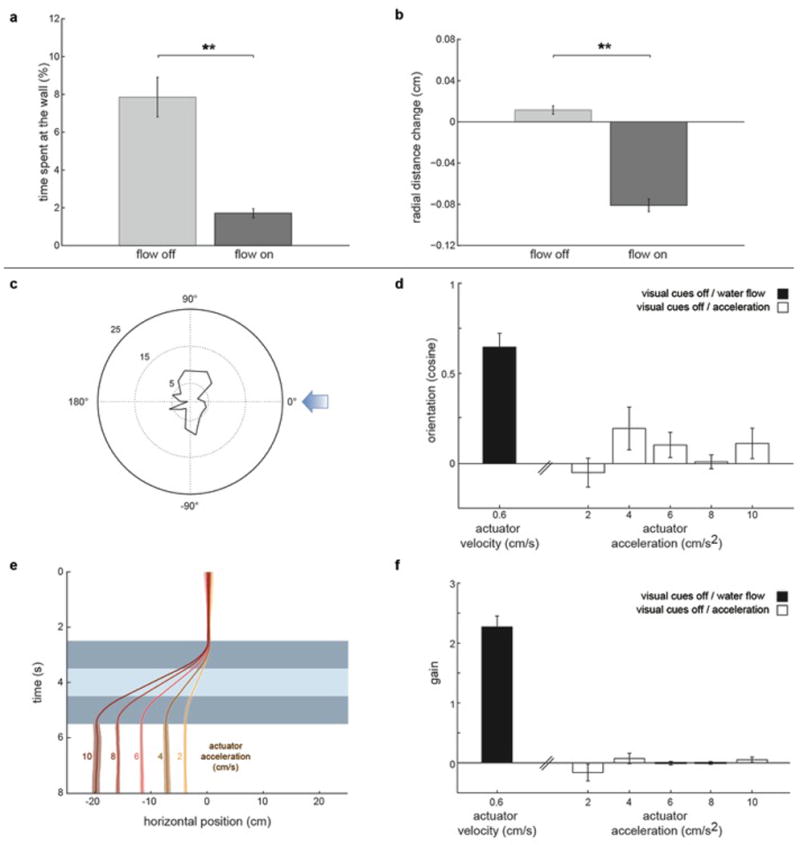
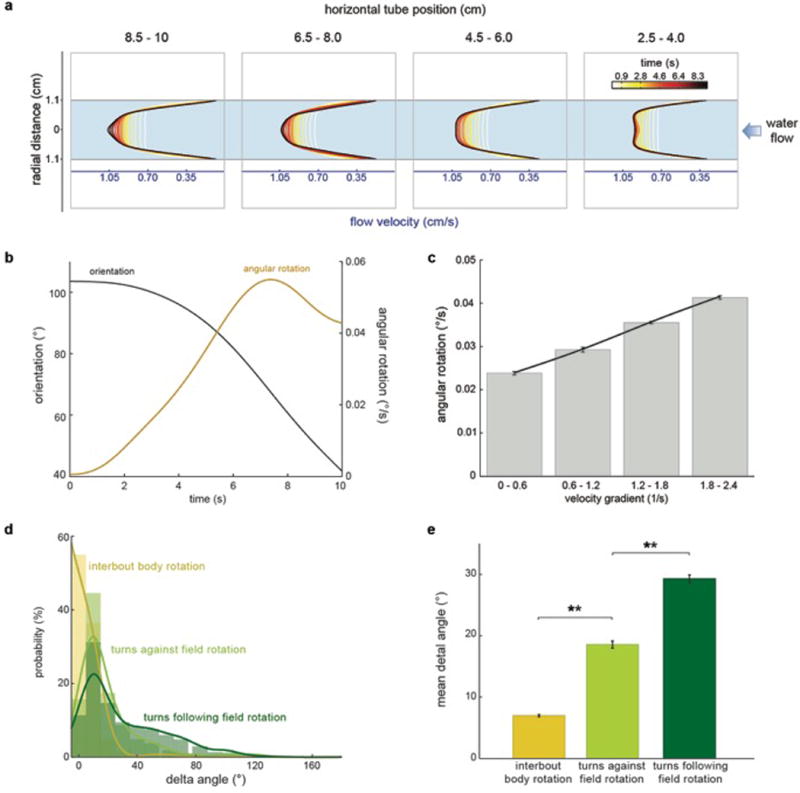
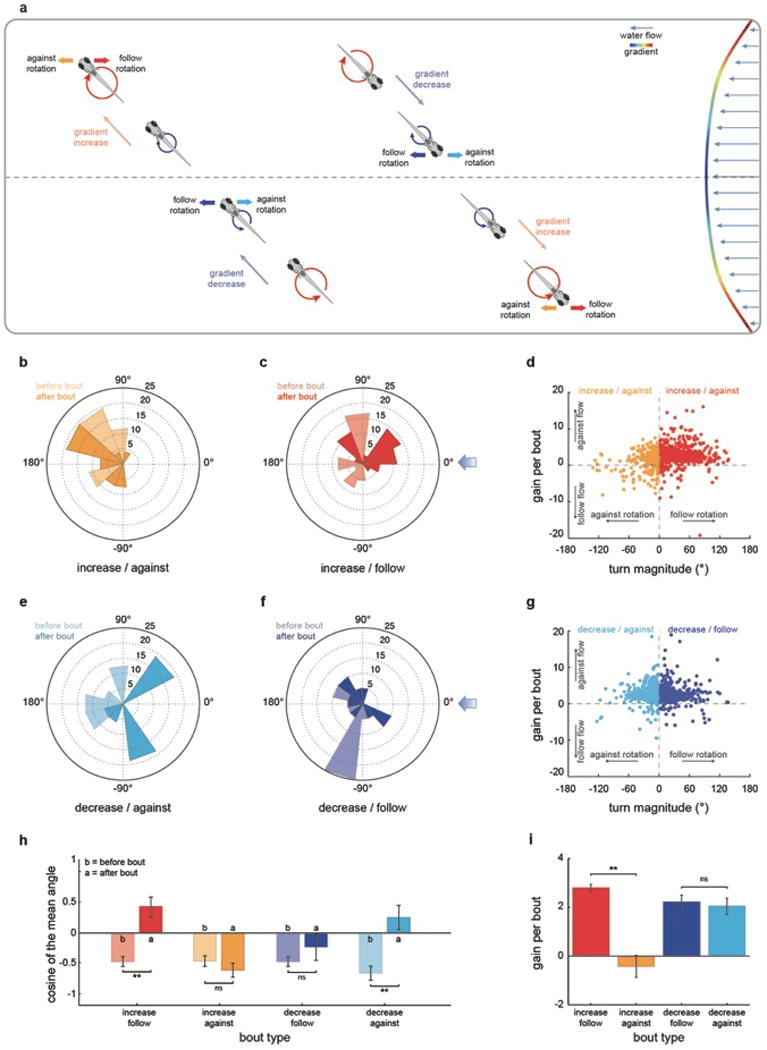
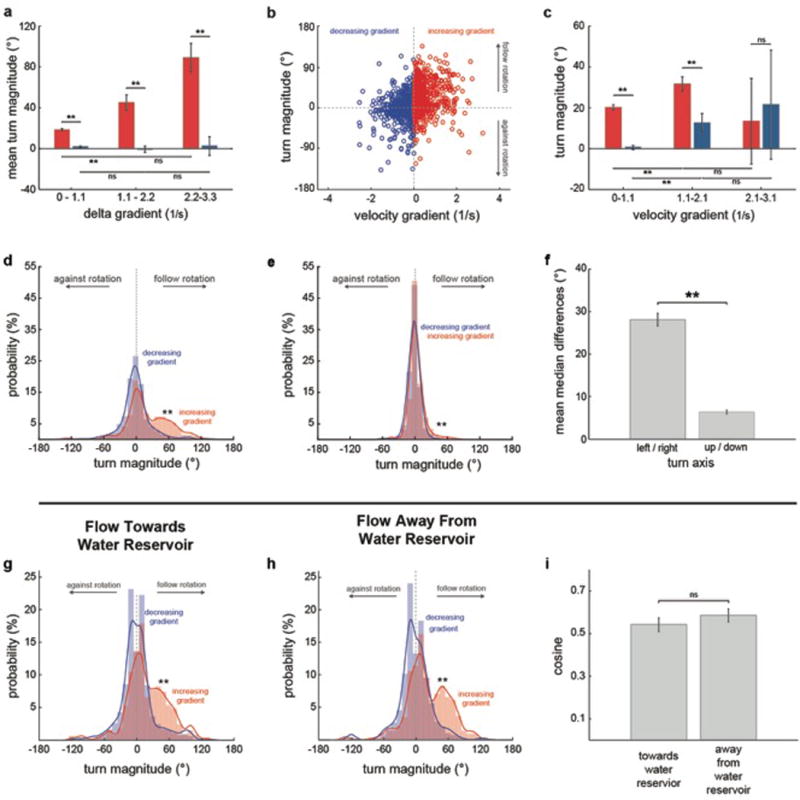
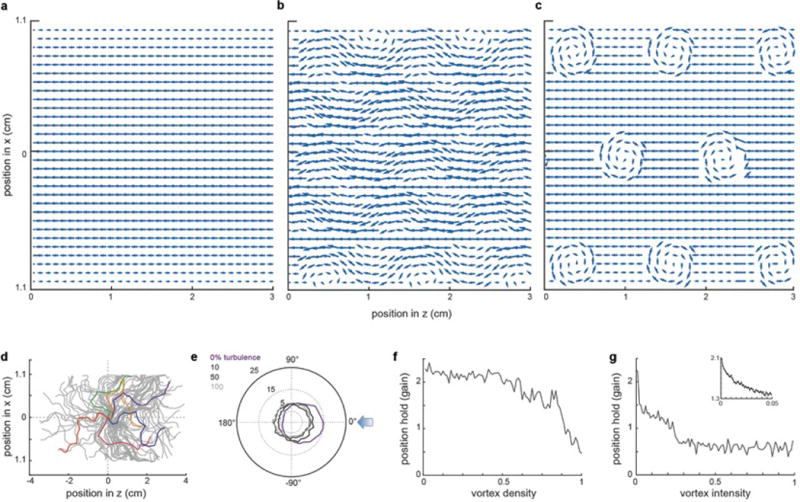
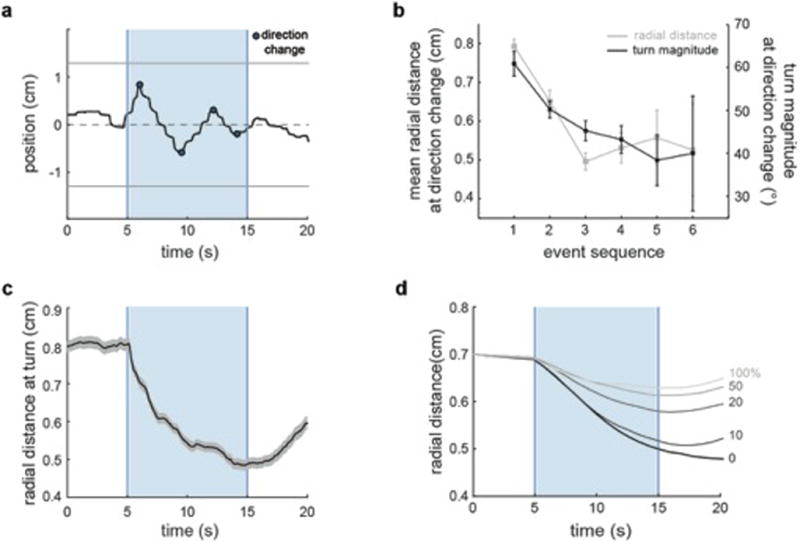
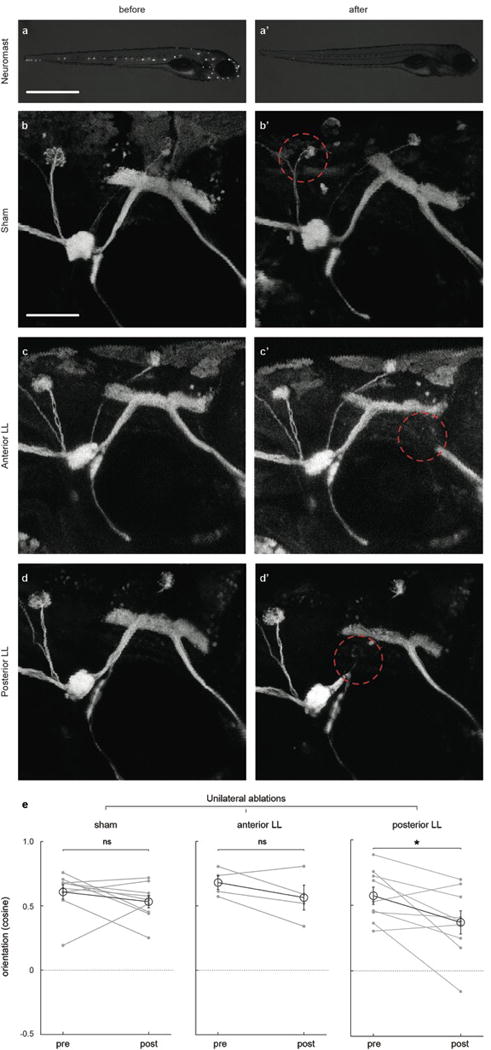
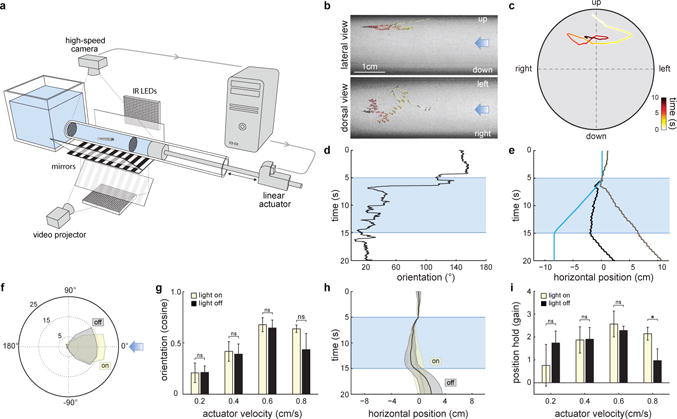
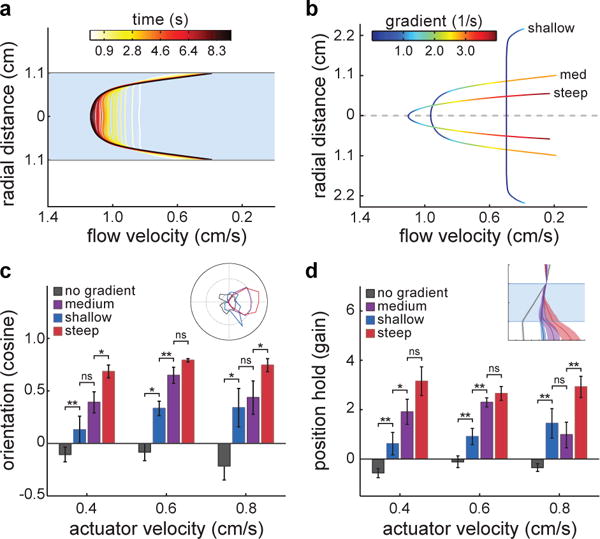
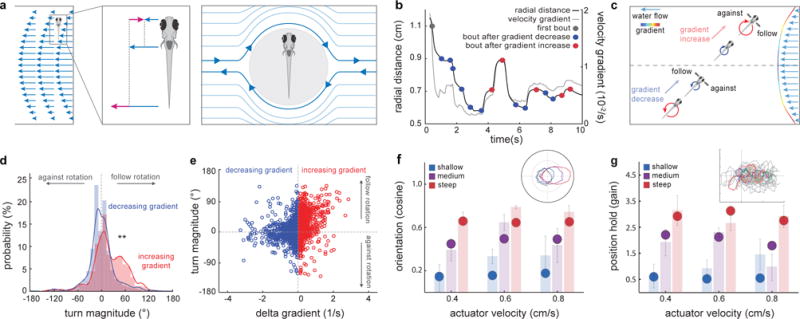
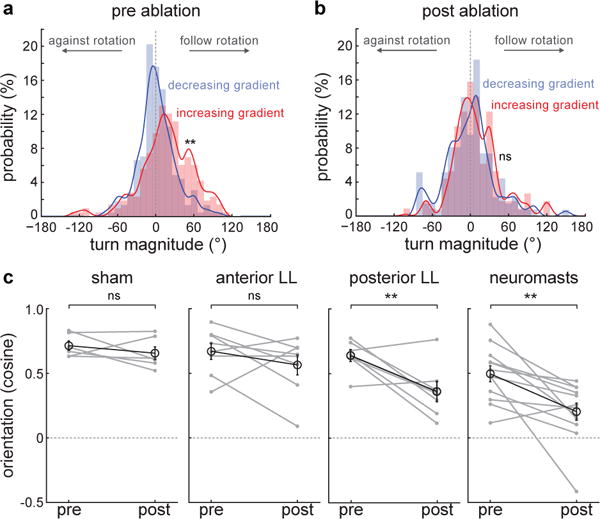
When flying or swimming, animals must adjust their own movement to compensate for displacements induced by the flow of the surrounding air or water. These flow-induced displacements can most easily be detected as visual whole-field motion with respect to the animal's frame of reference. Despite this, many aquatic animals consistently orient and swim against oncoming flows (a behaviour known as rheotaxis) even in the absence of visual cues. How animals achieve this task, and its underlying sensory basis, is still unknown. Here we show that, in the absence of visual information, larval zebrafish (Danio rerio) perform rheotaxis by using flow velocity gradients as navigational cues. We present behavioural data that support a novel algorithm based on such local velocity gradients that fish use to avoid getting dragged by flowing water. Specifically, we show that fish use their mechanosensory lateral line to first sense the curl (or vorticity) of the local velocity vector field to detect the presence of flow and, second, to measure its temporal change after swim bouts to deduce flow direction. These results reveal an elegant navigational strategy based on the sensing of flow velocity gradients and provide a comprehensive behavioural algorithm, also applicable for robotic design, that generalizes to a wide range of animal behaviours in moving fluids.
Conflict of interest statement
The authors declare no competing financial interests. Readers are welcome to comment on the online version of the paper.
Figures
Comment in
-
Biomechanics: How fish feel the flow.Nature. 2017 Jul 27;547(7664):406-407. doi: 10.1038/nature23096. Epub 2017 Jul 12. Nature. 2017. PMID: 28700572 No abstract available.
References
-
- Chapman JW, et al. Animal orientation strategies for movement in flows. Curr Biol. 2011;21:R861–870. - PubMed
-
- Lyon EP. On rheotropism I. Rheotropism in fishes. Am J Physiol. 1904;12:149–161.
-
- Dijkgraaf S. The functioning and significance of the lateral-line organs. Biol Rev Camb Philos Soc. 1963;38:51–105. - PubMed
-
- Montgomery J, Baker CF, Carton AG. The lateral line can mediate rheotaxis in fish. Nature. 1997;389:960–963.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
