

Systematic identification and characterization of regulatory elements derived from human endogenous retroviruses
- PMID: 28700586
- PMCID: PMC5529029
- DOI: 10.1371/journal.pgen.1006883
Systematic identification and characterization of regulatory elements derived from human endogenous retroviruses
Abstract
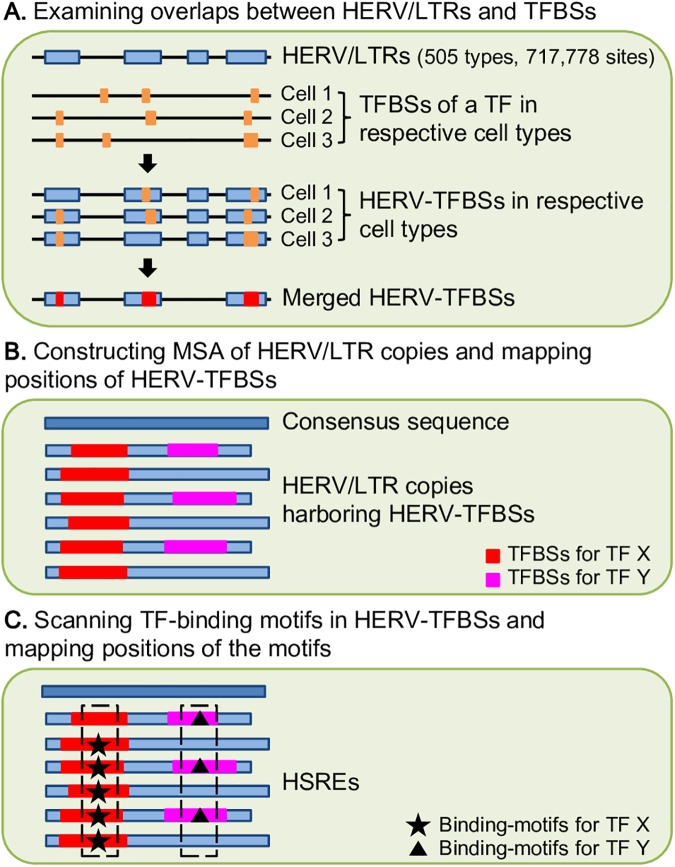
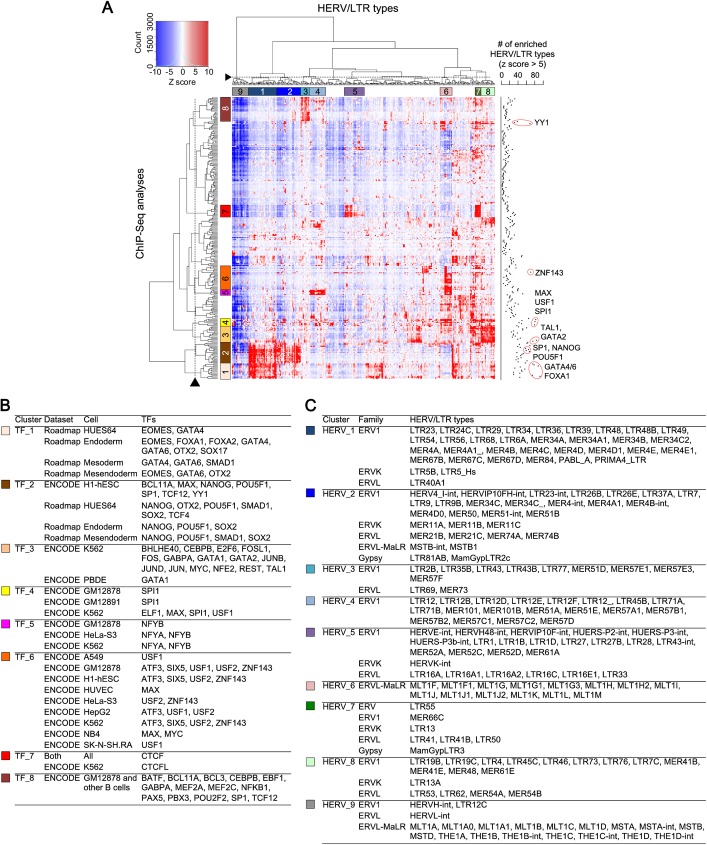
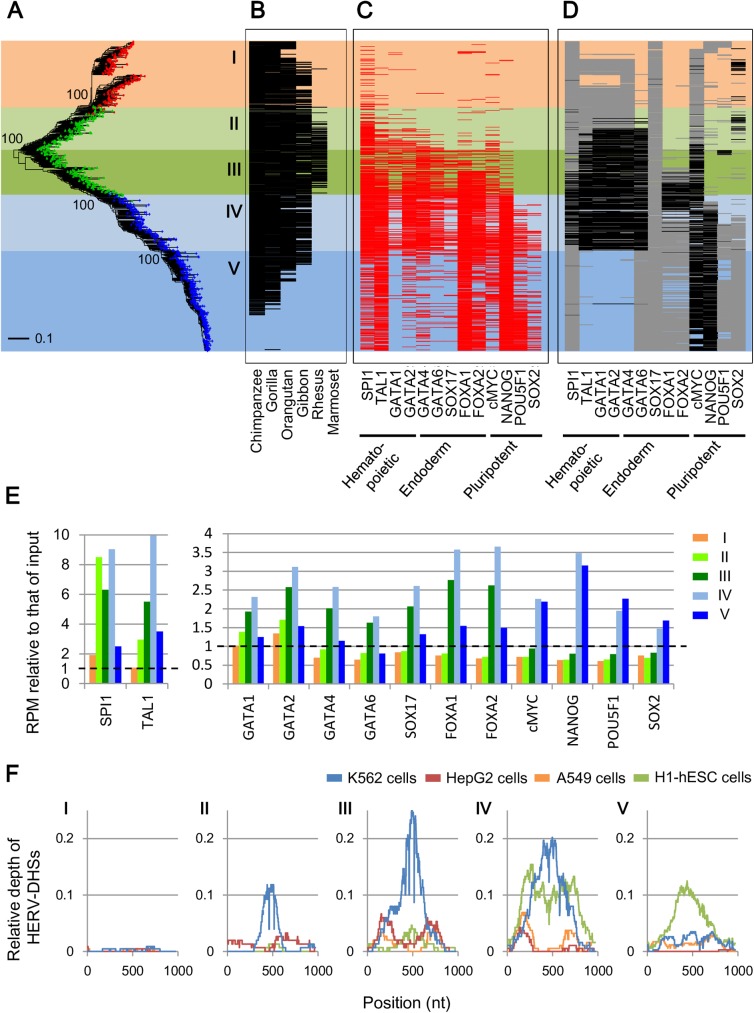
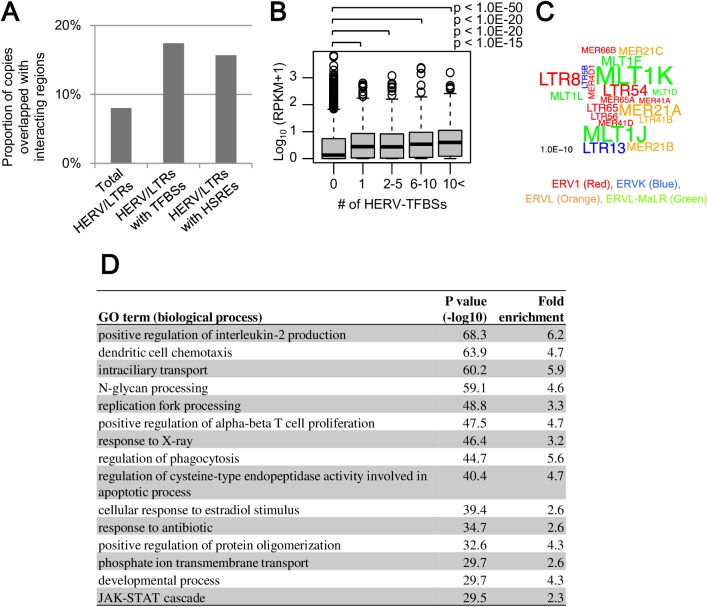
Human endogenous retroviruses (HERVs) and other long terminal repeat (LTR)-type retrotransposons (HERV/LTRs) have regulatory elements that possibly influence the transcription of host genes. We systematically identified and characterized these regulatory elements based on publicly available datasets of ChIP-Seq of 97 transcription factors (TFs) provided by ENCODE and Roadmap Epigenomics projects. We determined transcription factor-binding sites (TFBSs) using the ChIP-Seq datasets and identified TFBSs observed on HERV/LTR sequences (HERV-TFBSs). Overall, 794,972 HERV-TFBSs were identified. Subsequently, we identified "HERV/LTR-shared regulatory element (HSRE)," defined as a TF-binding motif in HERV-TFBSs, shared within a substantial fraction of a HERV/LTR type. HSREs could be an indication that the regulatory elements of HERV/LTRs are present before their insertions. We identified 2,201 HSREs, comprising specific associations of 354 HERV/LTRs and 84 TFs. Clustering analysis showed that HERV/LTRs can be grouped according to the TF binding patterns; HERV/LTR groups bounded to pluripotent TFs (e.g., SOX2, POU5F1, and NANOG), embryonic endoderm/mesendoderm TFs (e.g., GATA4/6, SOX17, and FOXA1/2), hematopoietic TFs (e.g., SPI1 (PU1), GATA1/2, and TAL1), and CTCF were identified. Regulatory elements of HERV/LTRs tended to locate nearby and/or interact three-dimensionally with the genes involved in immune responses, indicating that the regulatory elements play an important role in controlling the immune regulatory network. Further, we demonstrated subgroup-specific TF binding within LTR7, LTR5B, and LTR5_Hs, indicating that gains or losses of the regulatory elements occurred during genomic invasions of the HERV/LTRs. Finally, we constructed dbHERV-REs, an interactive database of HERV/LTR regulatory elements (http://herv-tfbs.com/). This study provides fundamental information in understanding the impact of HERV/LTRs on host transcription, and offers insights into the transcriptional modulation systems of HERV/LTRs and ancestral HERVs.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Similar articles
-
Molecular diversity and phenotypic pleiotropy of ancient genomic regulatory loci derived from human endogenous retrovirus type H (HERVH) promoter LTR7 and HERVK promoter LTR5_Hs and their contemporary impacts on pathophysiology of Modern Humans.Mol Genet Genomics. 2022 Nov;297(6):1711-1740. doi: 10.1007/s00438-022-01954-7. Epub 2022 Sep 19. Mol Genet Genomics. 2022. PMID: 36121513 Free PMC article.
-
p53 Binding Sites in Long Terminal Repeat 5Hs (LTR5Hs) of Human Endogenous Retrovirus K Family (HML-2 Subgroup) Play Important Roles in the Regulation of LTR5Hs Transcriptional Activity.Microbiol Spectr. 2022 Aug 31;10(4):e0048522. doi: 10.1128/spectrum.00485-22. Epub 2022 Jul 18. Microbiol Spectr. 2022. PMID: 35867400 Free PMC article.
-
Functional activity of HERV-K-T47D-related long terminal repeats.Virology. 2001 May 10;283(2):262-72. doi: 10.1006/viro.2001.0898. Virology. 2001. PMID: 11336551
-
The role of human endogenous retroviral long terminal repeat sequences in human cancer (Review).Int J Mol Med. 2013 Oct;32(4):755-62. doi: 10.3892/ijmm.2013.1460. Epub 2013 Jul 30. Int J Mol Med. 2013. PMID: 23900638 Review.
-
Utility of next-generation RNA-sequencing in identifying chimeric transcription involving human endogenous retroviruses.APMIS. 2016 Jan-Feb;124(1-2):127-39. doi: 10.1111/apm.12477. APMIS. 2016. PMID: 26818267 Review.
Cited by
-
The pluripotent stem cell-specific transcript ESRG is dispensable for human pluripotency.PLoS Genet. 2021 May 25;17(5):e1009587. doi: 10.1371/journal.pgen.1009587. eCollection 2021 May. PLoS Genet. 2021. PMID: 34033652 Free PMC article.
-
Genomics-Guided Drawing of Molecular and Pathophysiological Components of Malignant Regulatory Signatures Reveals a Pivotal Role in Human Diseases of Stem Cell-Associated Retroviral Sequences and Functionally-Active hESC Enhancers.Front Oncol. 2021 Mar 31;11:638363. doi: 10.3389/fonc.2021.638363. eCollection 2021. Front Oncol. 2021. PMID: 33869024 Free PMC article.
-
Genome-wide bioinformatic analyses predict key host and viral factors in SARS-CoV-2 pathogenesis.Commun Biol. 2021 May 17;4(1):590. doi: 10.1038/s42003-021-02095-0. Commun Biol. 2021. PMID: 34002013 Free PMC article.
-
Epigenomic analysis reveals prevalent contribution of transposable elements to cis-regulatory elements, tissue-specific expression, and alternative promoters in zebrafish.Genome Res. 2022 Jul;32(7):1424-1436. doi: 10.1101/gr.276052.121. Epub 2022 Jun 1. Genome Res. 2022. PMID: 35649578 Free PMC article.
-
Long Noncoding RNA HCP5, a Hybrid HLA Class I Endogenous Retroviral Gene: Structure, Expression, and Disease Associations.Cells. 2019 May 20;8(5):480. doi: 10.3390/cells8050480. Cells. 2019. PMID: 31137555 Free PMC article. Review.
References
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, et al. Initial sequencing and analysis of the human genome. Nature. 2001;409: 860–921. doi: 10.1038/35057062 - DOI - PubMed
-
- Hurst GD, Werren JH. The role of selfish genetic elements in eukaryotic evolution. Nat Rev Genet. 2001;2: 597–606. doi: 10.1038/35084545 - DOI - PubMed
-
- Feschotte C, Gilbert C. Endogenous viruses: insights into viral evolution and impact on host biology. Nat Rev Genet. 2012;13: 283–296. doi: 10.1038/nrg3199 - DOI - PubMed
-
- Mi S, Lee X, Li X, Veldman GM, Finnerty H, Racie L, et al. Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature. 2000;403: 785–789. doi: 10.1038/35001608 - DOI - PubMed
-
- Blaise S, de Parseval N, Benit L, Heidmann T. Genomewide screening for fusogenic human endogenous retrovirus envelopes identifies syncytin 2, a gene conserved on primate evolution. Proc Natl Acad Sci U S A. 2003;100: 13013–13018. doi: 10.1073/pnas.2132646100 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
