

Four simple rules that are sufficient to generate the mammalian blastocyst
- PMID: 28700688
- PMCID: PMC5507476
- DOI: 10.1371/journal.pbio.2000737
Four simple rules that are sufficient to generate the mammalian blastocyst
Abstract
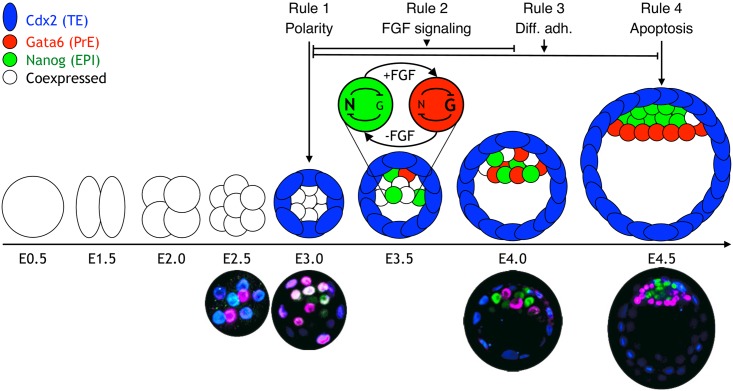
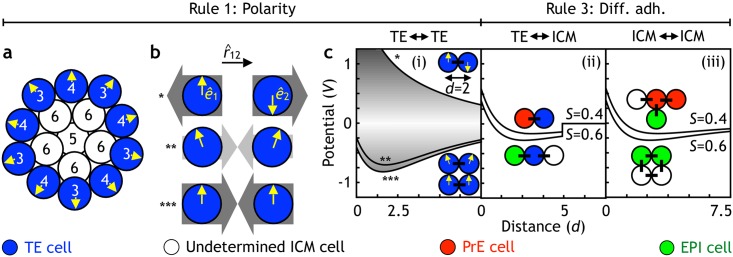
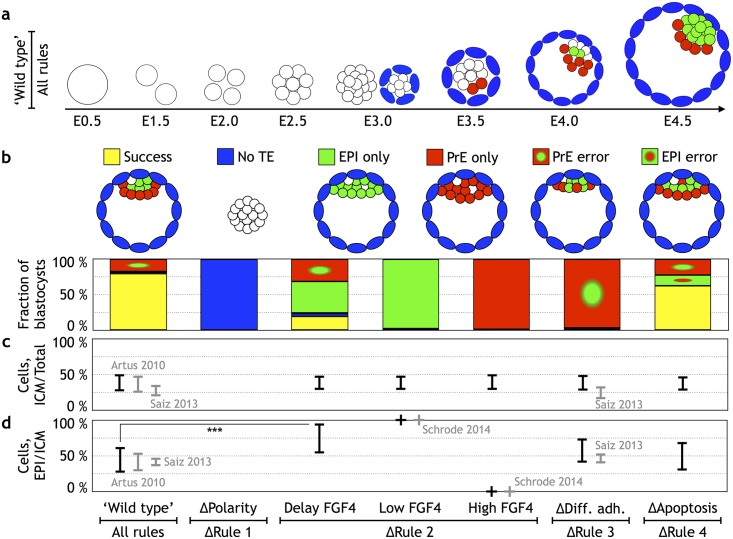
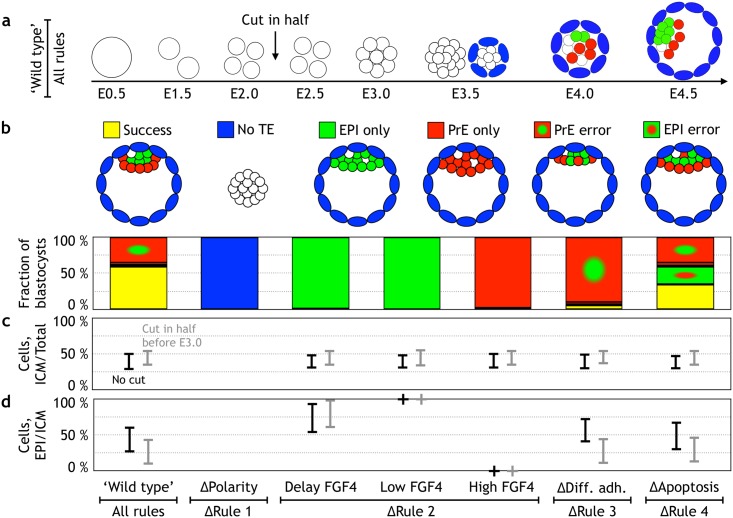
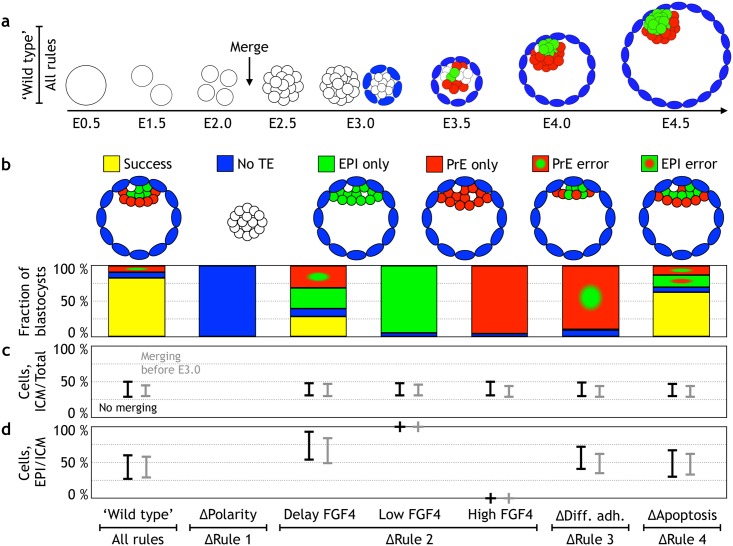
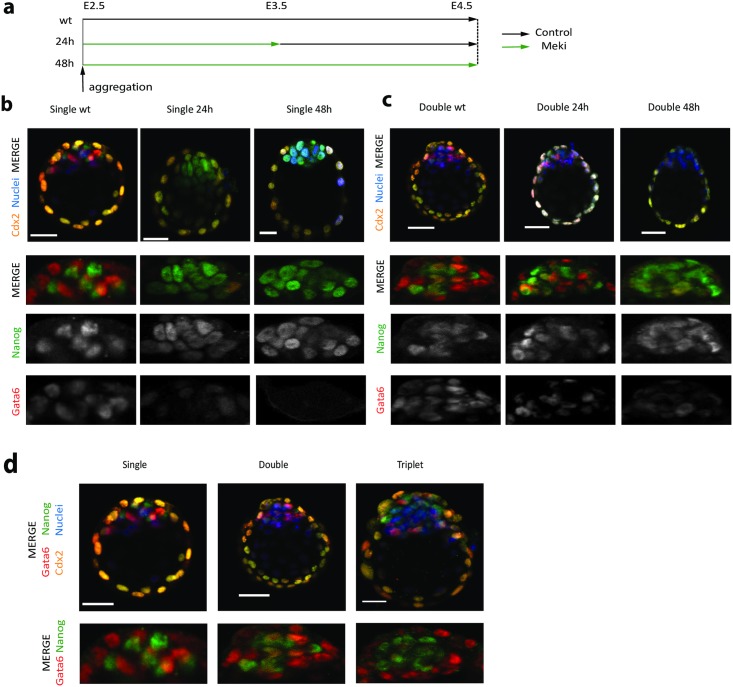
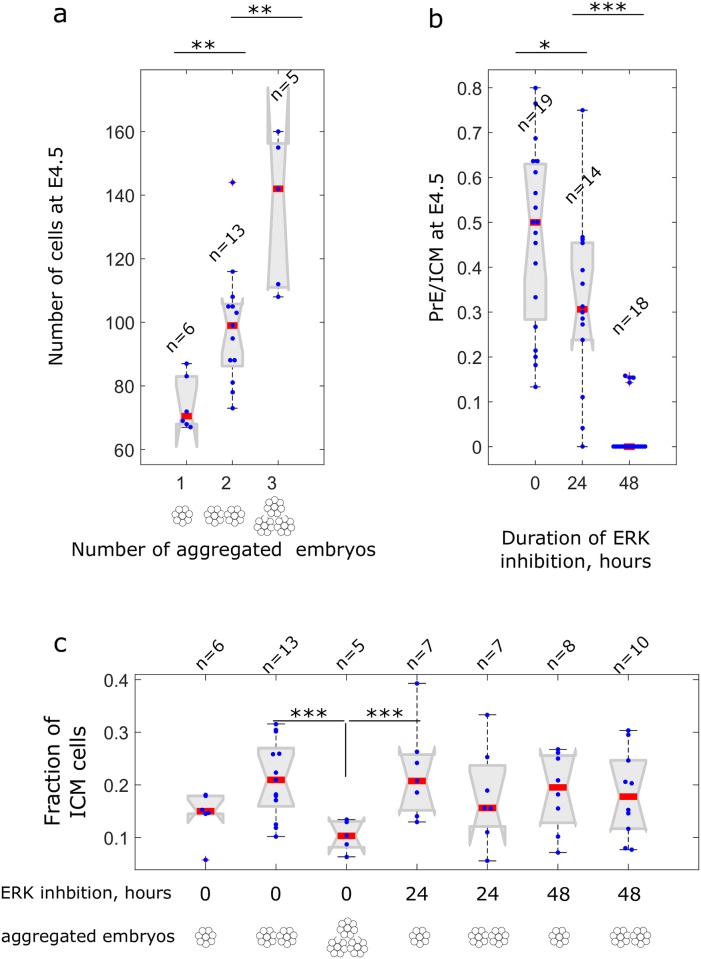
Early mammalian development is both highly regulative and self-organizing. It involves the interplay of cell position, predetermined gene regulatory networks, and environmental interactions to generate the physical arrangement of the blastocyst with precise timing. However, this process occurs in the absence of maternal information and in the presence of transcriptional stochasticity. How does the preimplantation embryo ensure robust, reproducible development in this context? It utilizes a versatile toolbox that includes complex intracellular networks coupled to cell-cell communication, segregation by differential adhesion, and apoptosis. Here, we ask whether a minimal set of developmental rules based on this toolbox is sufficient for successful blastocyst development, and to what extent these rules can explain mutant and experimental phenotypes. We implemented experimentally reported mechanisms for polarity, cell-cell signaling, adhesion, and apoptosis as a set of developmental rules in an agent-based in silico model of physically interacting cells. We find that this model quantitatively reproduces specific mutant phenotypes and provides an explanation for the emergence of heterogeneity without requiring any initial transcriptional variation. It also suggests that a fixed time point for the cells' competence of fibroblast growth factor (FGF)/extracellular signal-regulated kinase (ERK) sets an embryonic clock that enables certain scaling phenomena, a concept that we evaluate quantitatively by manipulating embryos in vitro. Based on these observations, we conclude that the minimal set of rules enables the embryo to experiment with stochastic gene expression and could provide the robustness necessary for the evolutionary diversification of the preimplantation gene regulatory network.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Cell fate regulation during preimplantation development: a view of adhesion-linked molecular interactions.Dev Biol. 2014 Nov 1;395(1):73-83. doi: 10.1016/j.ydbio.2014.08.028. Epub 2014 Aug 29. Dev Biol. 2014. PMID: 25176042 Review.
-
Gene Expression Noise Enhances Robust Organization of the Early Mammalian Blastocyst.PLoS Comput Biol. 2017 Jan 23;13(1):e1005320. doi: 10.1371/journal.pcbi.1005320. eCollection 2017 Jan. PLoS Comput Biol. 2017. PMID: 28114387 Free PMC article.
-
Signalling-modulated gene regulatory networks in early mammalian development.J Theor Biol. 2019 Feb 21;463:56-66. doi: 10.1016/j.jtbi.2018.12.008. Epub 2018 Dec 10. J Theor Biol. 2019. PMID: 30543809
-
Signaling pathways and preimplantation development of mammalian embryos.FEBS J. 2007 Sep;274(17):4349-59. doi: 10.1111/j.1742-4658.2007.05980.x. Epub 2007 Aug 3. FEBS J. 2007. PMID: 17680809 Review.
-
Simulating the mammalian blastocyst--molecular and mechanical interactions pattern the embryo.PLoS Comput Biol. 2011 May;7(5):e1001128. doi: 10.1371/journal.pcbi.1001128. Epub 2011 May 5. PLoS Comput Biol. 2011. PMID: 21573197 Free PMC article.
Cited by
-
The primitive endoderm supports lineage plasticity to enable regulative development.Cell. 2024 Jul 25;187(15):4010-4029.e16. doi: 10.1016/j.cell.2024.05.051. Epub 2024 Jun 24. Cell. 2024. PMID: 38917790 Free PMC article.
-
Generative model for the first cell fate bifurcation in mammalian development.bioRxiv [Preprint]. 2025 Feb 25:2025.02.24.639895. doi: 10.1101/2025.02.24.639895. bioRxiv. 2025. Update in: Development. 2025 Aug 05:dev.204717. doi: 10.1242/dev.204717. PMID: 40060535 Free PMC article. Updated. Preprint.
-
Lineage tracing identifies heterogeneous hepatoblast contribution to cell lineages and postembryonic organ growth dynamics.PLoS Biol. 2023 Oct 4;21(10):e3002315. doi: 10.1371/journal.pbio.3002315. eCollection 2023 Oct. PLoS Biol. 2023. PMID: 37792696 Free PMC article.
-
Effective mechanical potential of cell-cell interaction explains three-dimensional morphologies during early embryogenesis.PLoS Comput Biol. 2023 Aug 7;19(8):e1011306. doi: 10.1371/journal.pcbi.1011306. eCollection 2023 Aug. PLoS Comput Biol. 2023. PMID: 37549166 Free PMC article.
-
Biophysical models of early mammalian embryogenesis.Stem Cell Reports. 2023 Jan 10;18(1):26-46. doi: 10.1016/j.stemcr.2022.11.021. Stem Cell Reports. 2023. PMID: 36630902 Free PMC article. Review.
References
-
- Cockburn K, Rossant J. Making the blastocyst: lessons from the mouse. J Clin Invest. 2010;120: 995–1003. doi: 10.1172/JCI41229 - DOI - PMC - PubMed
-
- Wicklow E, Blij S, Frum T, Hirate Y, Lang RA, Sasaki H, et al. HIPPO Pathway Members Restrict SOX2 to the Inner Cell Mass Where It Promotes ICM Fates in the Mouse Blastocyst. PLoS Genet. 2014;10 doi: 10.1371/journal.pgen.1004618 - DOI - PMC - PubMed
-
- Chazaud C, Yamanaka Y, Pawson T, Rossant J. Early Lineage Segregation between Epiblast and Primitive Endoderm in Mouse Blastocysts through the Grb2-MAPK Pathway. Dev Cell. 2006;10: 615–24. doi: 10.1016/j.devcel.2006.02.020 - DOI - PubMed
-
- Plusa B, Piliszek A, Frankenberg S, Artus J, Hadjantonakis A-K. Distinct sequential cell behaviours direct primitive endoderm formation in the mouse blastocyst. Development. 2008;135: 3081–91. doi: 10.1242/dev.021519 - DOI - PMC - PubMed
-
- Van De Velde H, Cauffman G, Tournaye H, Devroey P, Liebaers I. The four blastomeres of a 4-cell stage human embryo are able to develop individually into blastocysts with inner cell mass and trophectoderm. Hum Reprod. 2008;23: 1742–1747. doi: 10.1093/humrep/den190 - DOI - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
