

Interleukin-3 enhances the migration of human mesenchymal stem cells by regulating expression of CXCR4
- PMID: 28705238
- PMCID: PMC5512829
- DOI: 10.1186/s13287-017-0618-y
Interleukin-3 enhances the migration of human mesenchymal stem cells by regulating expression of CXCR4
Abstract
Background: Mesenchymal stem cells (MSCs) represent an important source for cell therapy in regenerative medicine. MSCs have shown promising results for repair of damaged tissues in various degenerative diseases in animal models and also in human clinical trials. However, little is known about the factors that could enhance the migration and tissue-specific engraftment of exogenously infused MSCs for successful regenerative cell therapy. Previously, we have reported that interleukin-3 (IL-3) prevents bone and cartilage damage in animal models of rheumatoid arthritis and osteoarthritis. Also, IL-3 promotes the differentiation of human MSCs into functional osteoblasts and increases their in-vivo bone regenerative potential in immunocompromised mice. However, the role of IL-3 in migration of MSCs is not yet known. In the present study, we investigated the role of IL-3 in migration of human MSCs under both in-vitro and in-vivo conditions.
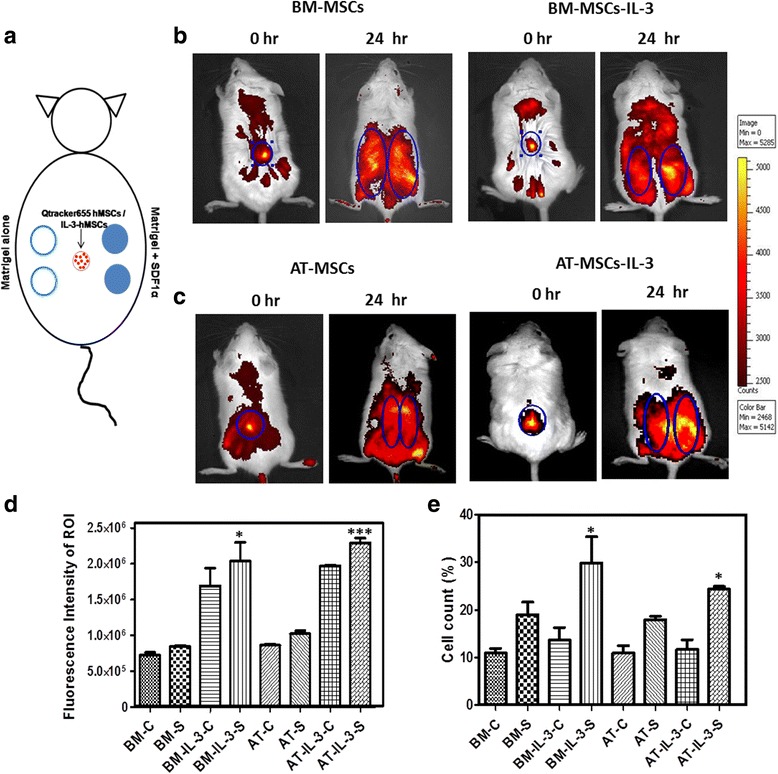
Methods: MSCs isolated from human bone marrow, adipose and gingival tissues were used for in-vitro cell migration, motility and wound healing assays in the presence or absence of IL-3. The effect of IL-3 preconditioning on expression of chemokine receptors and integrins was examined by flow cytometry and real-time PCR. The in-vivo migration of IL-3-preconditioned MSCs was investigated using a subcutaneous matrigel-releasing stromal cell-derived factor-1 alpha (SDF-1α) model in immunocompromised mice.
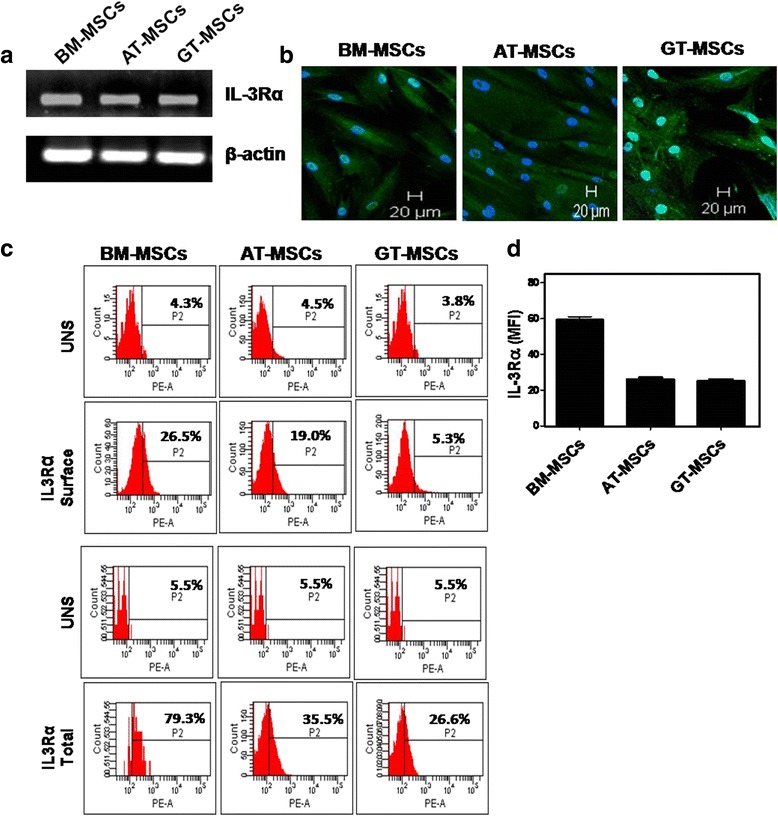
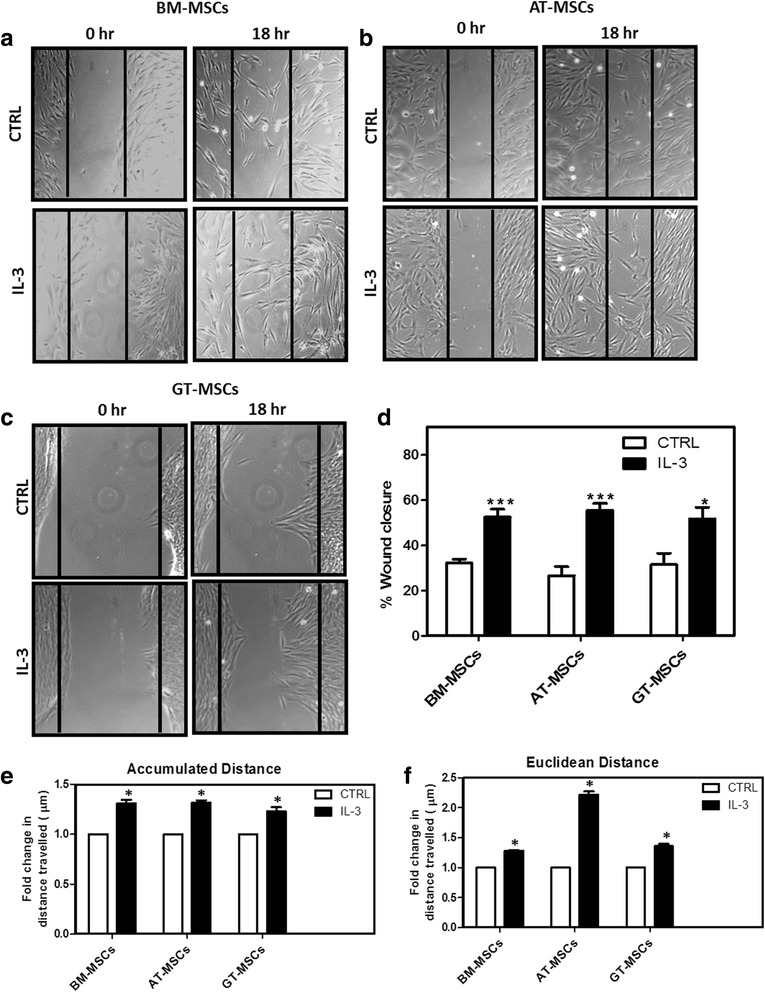
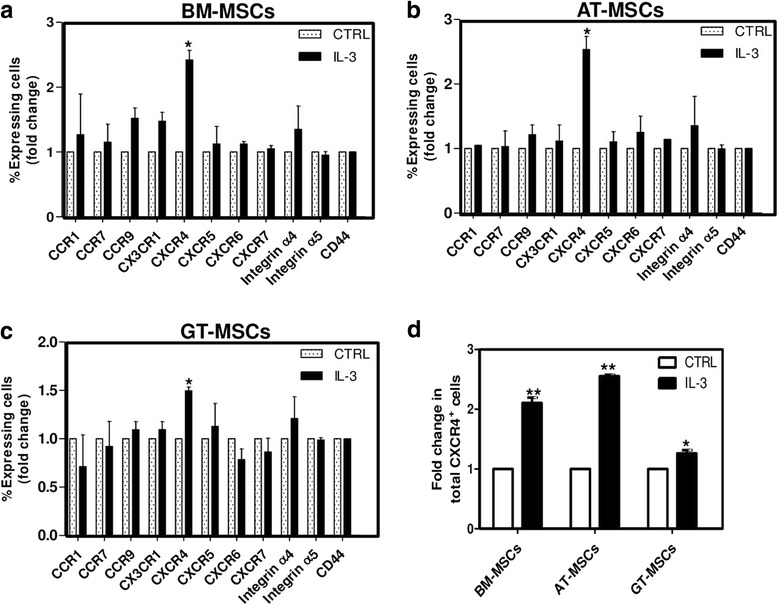
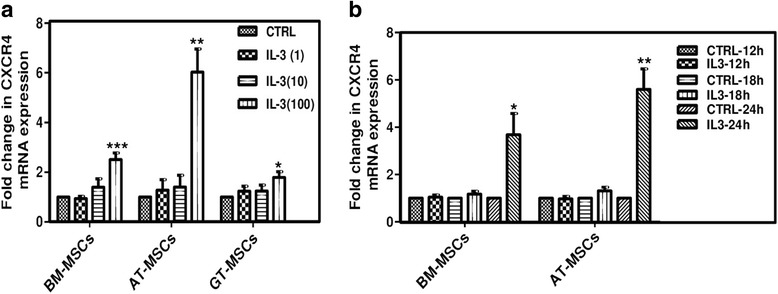
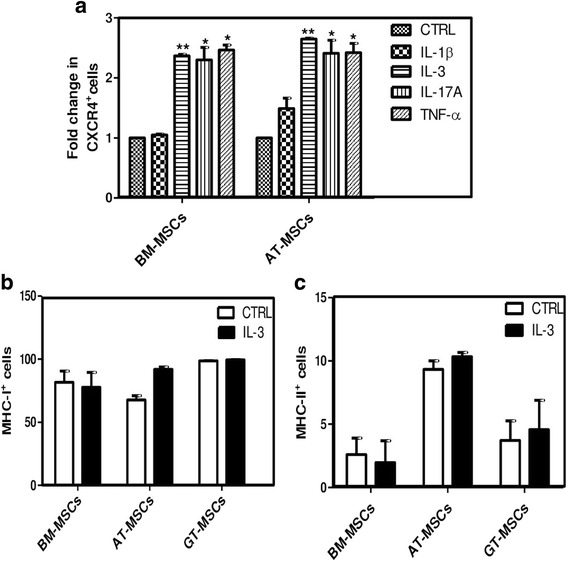
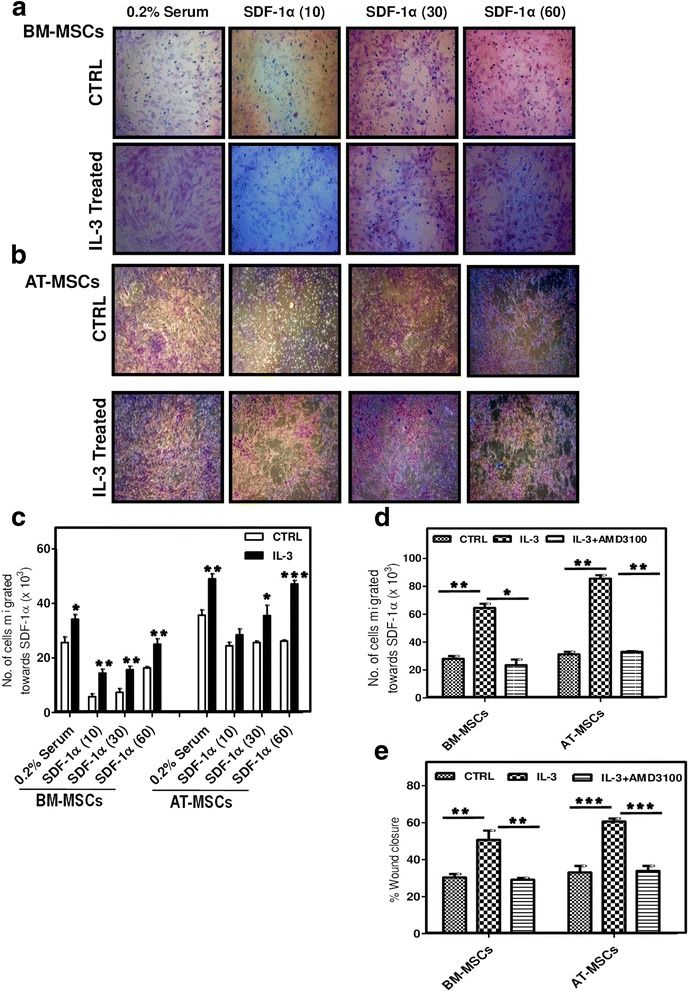
Results: We observed that human MSCs isolated from all three sources express IL-3 receptor-α (IL-3Rα) both at gene and protein levels. IL-3 significantly enhances in-vitro migration, motility and wound healing abilities of MSCs. Moreover, IL-3 preconditioning upregulates expression of chemokine (C-X-C motif) receptor 4 (CXCR4) on MSCs, which leads to increased migration of cells towards SDF-1α. Furthermore, CXCR4 antagonist AMD3100 decreases the migration of IL-3-treated MSCs towards SDF-1α. Importantly, IL-3 also induces in-vivo migration of MSCs towards subcutaneously implanted matrigel-releasing-SDF-1α in immunocompromised mice.
Conclusions: The present study demonstrates for the first time that IL-3 has an important role in enhancing the migration of human MSCs through regulation of the CXCR4/SDF-1α axis. These findings suggest a potential role of IL-3 in improving the efficacy of MSCs in regenerative cell therapy.
Keywords: CXCR4; Cell migration; Interleukin-3; Mesenchymal stem cells; SDF-1α.
Conflict of interest statement
Authors’ information
Not applicable.
Ethics approval and consent to participate
Protocols for animal experiments were approved by the Institutional Animal Ethical Committee (IAEC) of the National Centre for Cell Science, Pune, India (EAF/2014/B-237).
Human bone marrow, adipose tissue and gingival tissue samples were collected from local hospitals after obtaining informed consent with the compliance of the Institutional Ethical Committee (IEC) and the Institutional Committee for Stem Cell Research (IC-SCR) of the National Centre for Cell Science, Pune, India (NCCS/IC-SCR/2013/7).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Barbash IM, Chouraqui P, Baron J, Feinberg MS, Etzion S, Tessone A, et al. Systemic delivery of bone marrow-derived mesenchymal stem cells to the infarcted myocardium: feasibility, cell migration, and body distribution. Circulation. 2003;108:863–68. doi: 10.1161/01.CIR.0000084828.50310.6A. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
