

Vector control with driving Y chromosomes: modelling the evolution of resistance
- PMID: 28705249
- PMCID: PMC5513332
- DOI: 10.1186/s12936-017-1932-7
Vector control with driving Y chromosomes: modelling the evolution of resistance
Abstract
Background: The introduction of new malaria control interventions has often led to the evolution of resistance, both of the parasite to new drugs and of the mosquito vector to new insecticides, compromising the efficacy of the interventions. Recent progress in molecular and population biology raises the possibility of new genetic-based interventions, and the potential for resistance to evolve against these should be considered. Here, population modelling is used to determine the main factors affecting the likelihood that resistance will evolve against a synthetic, nuclease-based driving Y chromosome that produces a male-biased sex ratio.
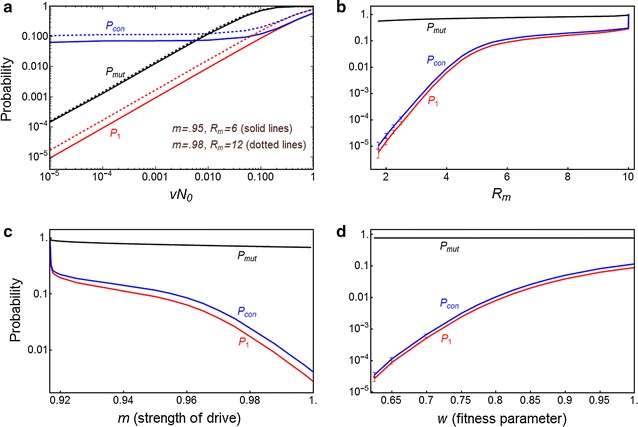
Methods: A combination of deterministic differential equation models and stochastic analyses involving branching processes and Gillespie simulations is utilized to assess the probability that resistance evolves against a driving Y that otherwise is strong enough to eliminate the target population. The model considers resistance due to changes at the target site such that they are no longer cleaved by the nuclease, and due to trans-acting autosomal suppressor alleles.
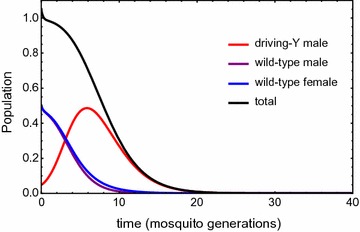
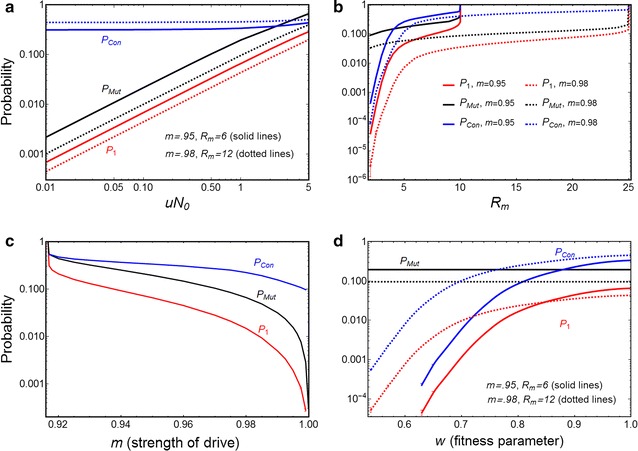
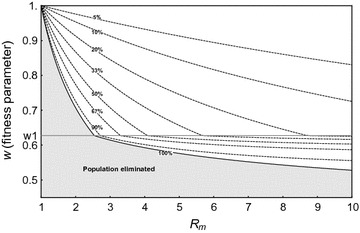
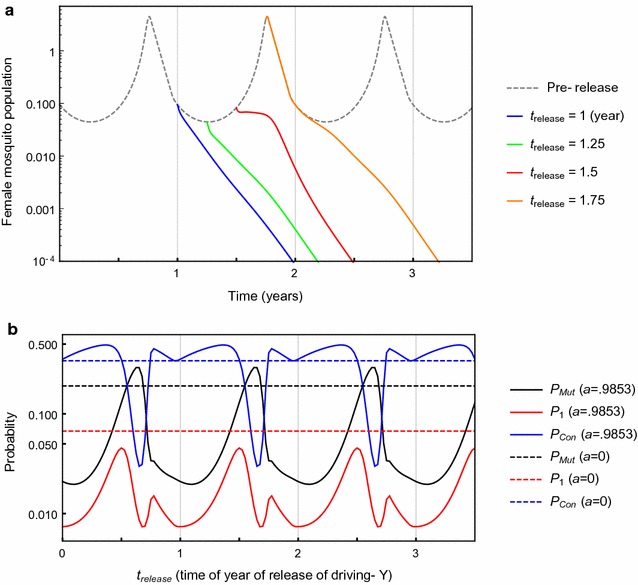
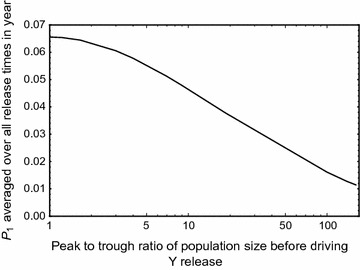
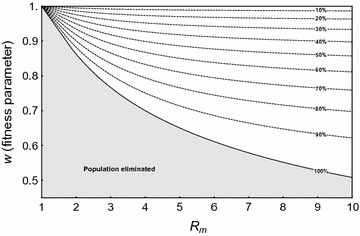
Results: The probability that resistance evolves increases with the mutation rate and the intrinsic rate of increase of the population, and decreases with the strength of drive and any pleiotropic fitness costs of the resistant allele. In seasonally varying environments, the time of release can also affect the probability of resistance evolving. Trans-acting suppressor alleles are more likely to suffer stochastic loss at low frequencies than target site resistant alleles.
Conclusions: As with any other intervention, there is a risk that resistance will evolve to new genetic approaches to vector control, and steps should be taken to minimize this probability. Two design features that should help in this regard are to reduce the rate at which resistant mutations arise, and to target sequences such that if they do arise, they impose a significant fitness cost on the mosquito.
Keywords: Anopheles gambiae; Branching process; Driving Y chromosome; Gene drive; Malaria; Resistance; Vector control.
Figures
References
-
- Barnes KG, Weedall GD, Ndula M, Irving H, Mzihalowa T, Hemingway J, et al. Genomic footprints of selective sweeps from metabolic resistance to pyrethroids in African malaria vectors are driven by scale up of insecticide-based vector control. PLoS Genet. 2017;13:e1006539. doi: 10.1371/journal.pgen.1006539. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
