

Hyperactive FOXO1 results in lack of tip stalk identity and deficient microvascular regeneration during kidney injury
- PMID: 28711779
- PMCID: PMC5567800
- DOI: 10.1016/j.biomaterials.2017.07.010
Hyperactive FOXO1 results in lack of tip stalk identity and deficient microvascular regeneration during kidney injury
Abstract
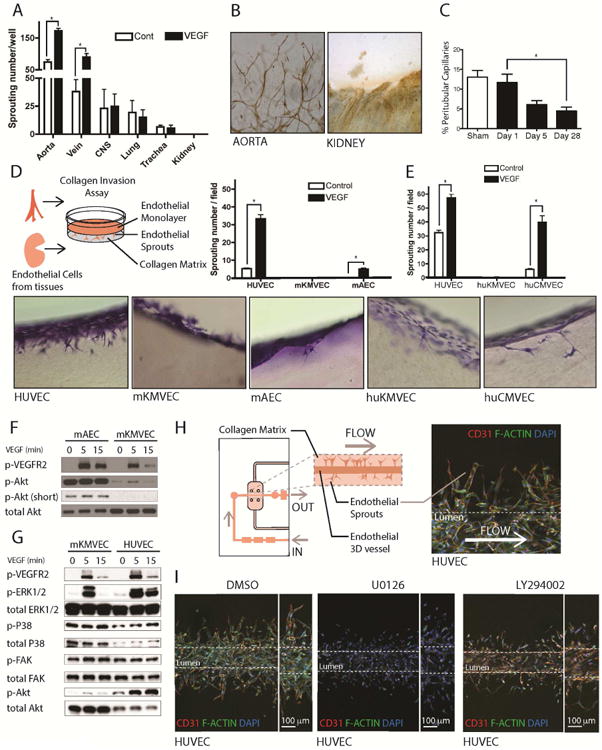
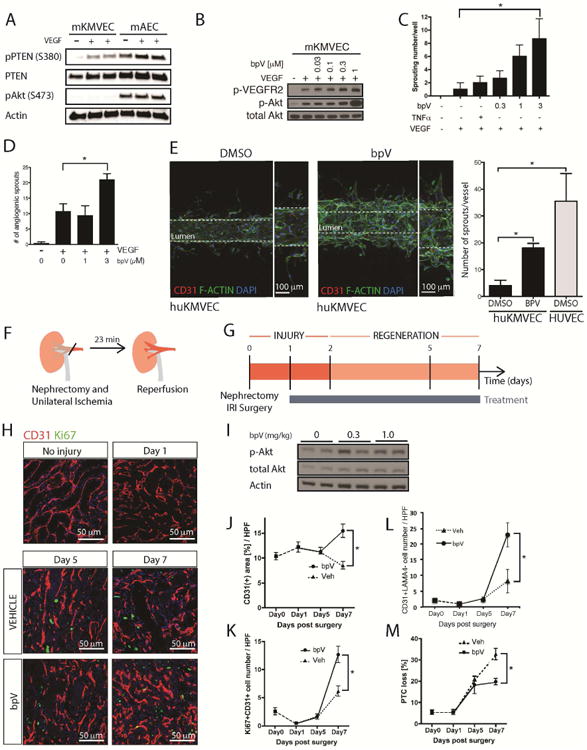
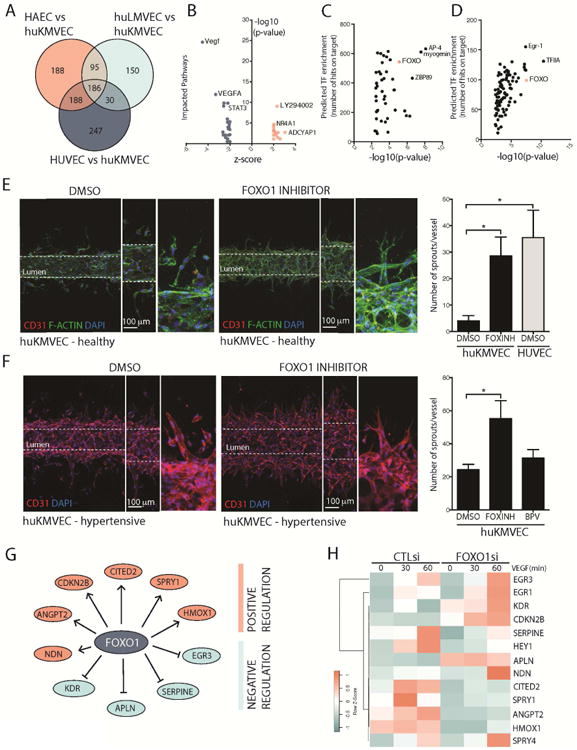
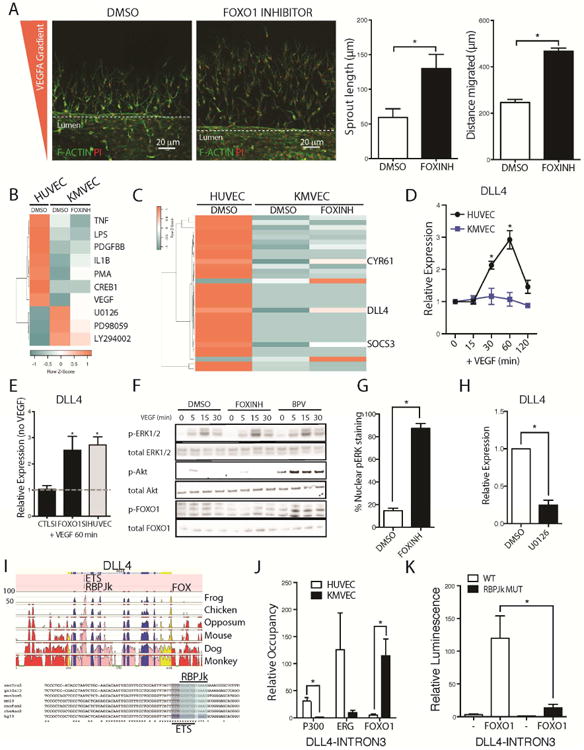
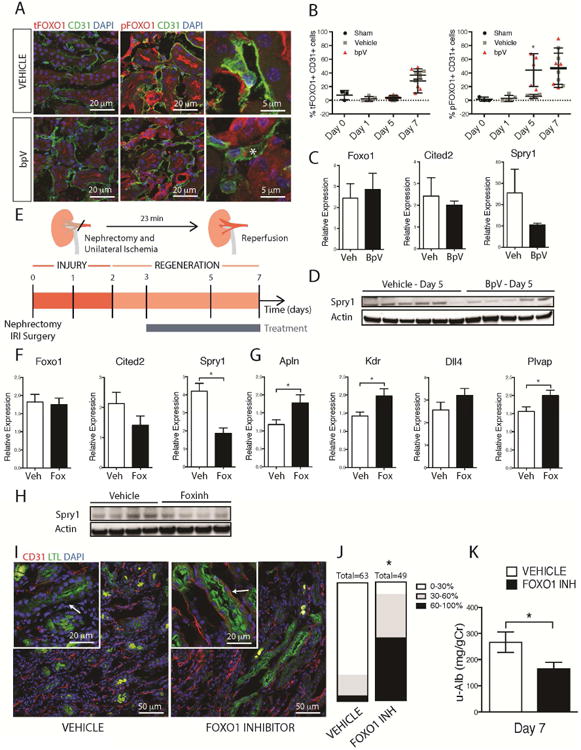
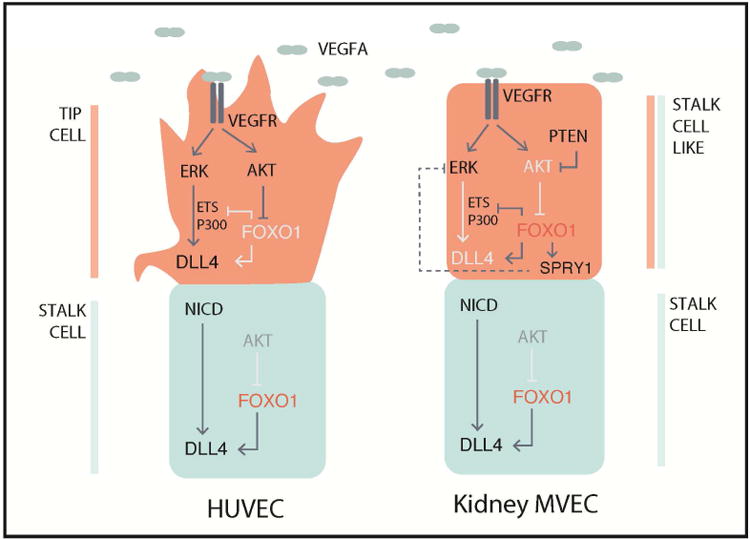
Loss of the microvascular (MV) network results in tissue ischemia, loss of tissue function, and is a hallmark of chronic diseases. The incorporation of a functional vascular network with that of the host remains a challenge to utilizing engineered tissues in clinically relevant therapies. We showed that vascular-bed-specific endothelial cells (ECs) exhibit differing angiogenic capacities, with kidney microvascular endothelial cells (MVECs) being the most deficient, and sought to explore the underlying mechanism. Constitutive activation of the phosphatase PTEN in kidney MVECs resulted in impaired PI3K/AKT activity in response to vascular endothelial growth factor (VEGF). Suppression of PTEN in vivo resulted in microvascular regeneration, but was insufficient to improve tissue function. Promoter analysis of the differentially regulated genes in KMVECs suggests that the transcription factor FOXO1 is highly active and RNAseq analysis revealed that hyperactive FOXO1 inhibits VEGF-Notch-dependent tip-cell formation by direct and indirect inhibition of DLL4 expression in response to VEGF. Inhibition of FOXO1 enhanced angiogenesis in human bio-engineered capillaries, and resulted in microvascular regeneration and improved function in mouse models of injury-repair.
Keywords: Angiogenesis; FOXO1; Microfluidics; VEGF; Vascular rarefaction; Vascular regeneration.
Copyright © 2017 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Figures
References
-
- Ye L, et al. Transplantation of nanoparticle transfected skeletal myoblasts overexpressing vascular endothelial growth factor-165 for cardiac repair. Circulation. 2007;116(11 Suppl):I113–20. - PubMed
-
- Kolesky DB, et al. 3D bioprinting of vascularized, heterogeneous cell-laden tissue constructs. Adv Mater. 2014;26(19):3124–30. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
