

Immune dysfunctionality of replicative senescent mesenchymal stromal cells is corrected by IFNγ priming
- PMID: 28713871
- PMCID: PMC5507374
- DOI: 10.1182/bloodadvances.2017006205
Immune dysfunctionality of replicative senescent mesenchymal stromal cells is corrected by IFNγ priming
Abstract
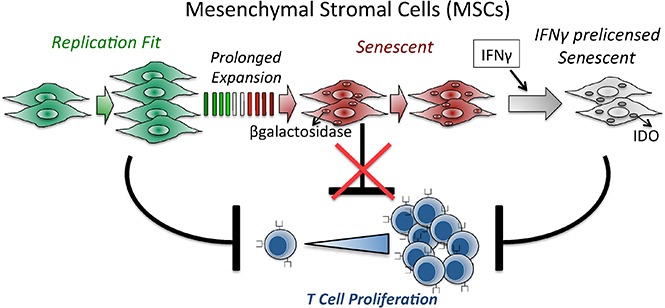
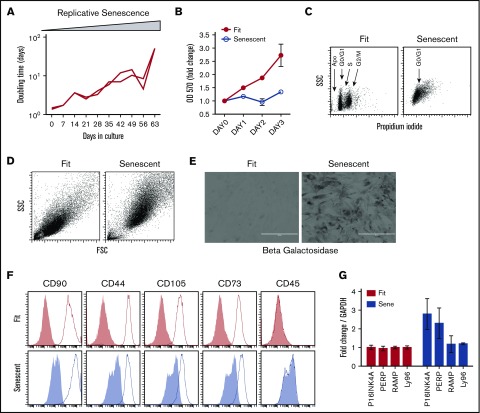
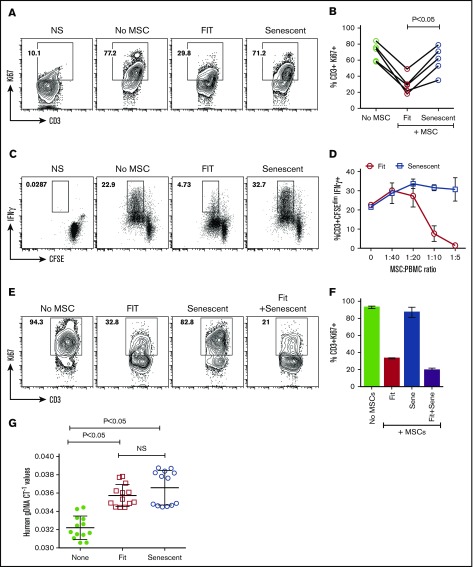
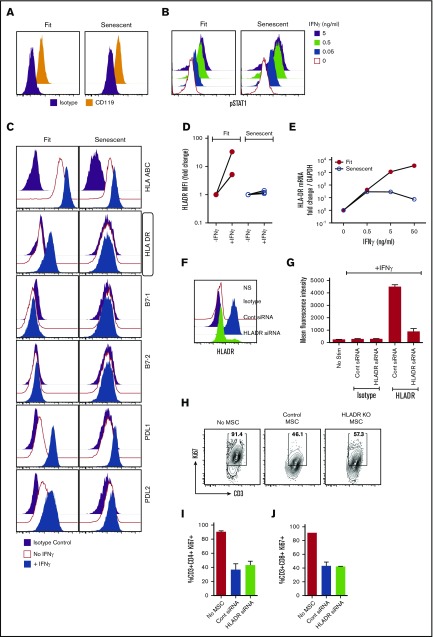
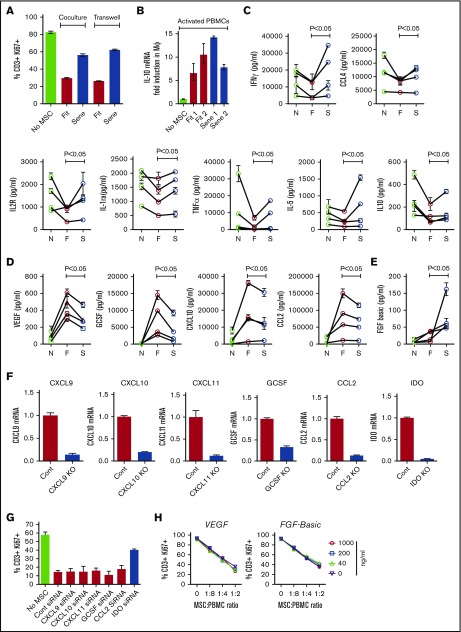
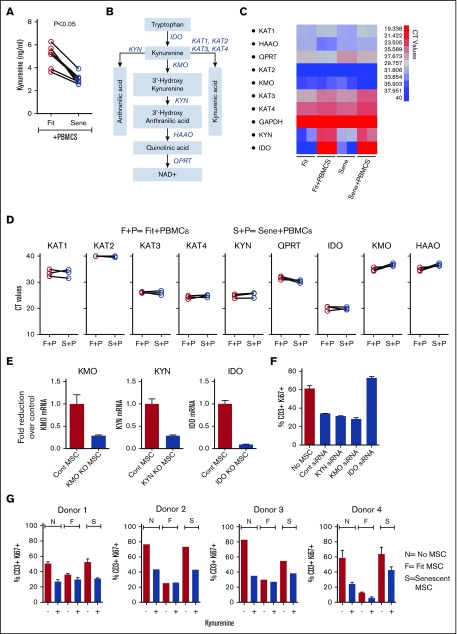
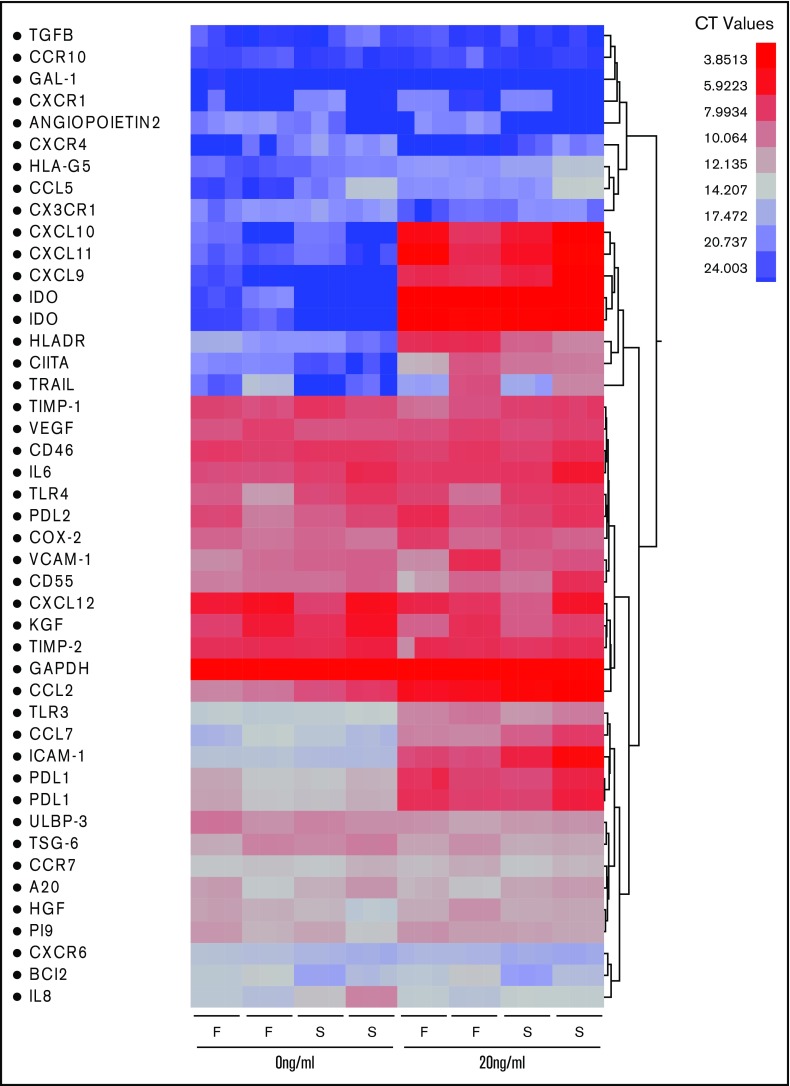
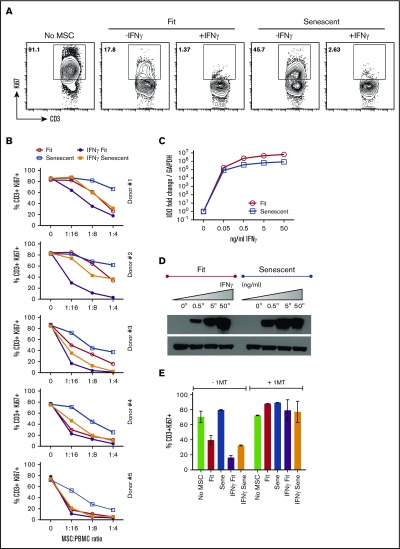
Industrial-scale expansion of mesenchymal stromal cells (MSCs) is often used in clinical trials, and the effect of replicative senescence on MSC functionality is of mechanistic interest. Senescent MSCs exhibit cell-cycle arrest, cellular hypertrophy, and express the senescent marker β-galactosidase. Although both fit and senescent MSCs display intact lung-homing properties in vivo, senescent MSCs acquire a significant defect in inhibiting T-cell proliferation and cytokine secretion in vitro. IFNγ does not upregulate HLA-DR on senescent MSCs, whereas its silencing did not reverse fit MSCs' immunosuppressive properties. Secretome analysis of MSC and activated peripheral blood mononuclear cell coculture demonstrate that senescent MSCs are significantly defective in up (vascular endothelial growth factor [VEGF], granulocyte colony-stimulating factor [GCSF], CXCL10, CCL2) or down (IL-1ra, IFNγ, IL-2r, CCL4, tumor necrosis factor-α, IL-5) regulating cytokines/chemokines. Unlike indoleamine 2,3 dioxygenase (IDO), silencing of CXCL9, CXCL10, CXCL11, GCSF, CCL2, and exogenous addition of VEGF, fibroblast growth factor-basic do not modulate MSCs' immunosuppressive properties. Kynurenine levels were downregulated in senescent MSC cocultures compared with fit MSC counterparts, and exogenous addition of kynurenine inhibits T-cell proliferation in the presence of senescent MSCs. IFNγ prelicensing activated several immunomodulatory genes including IDO in fit and senescent MSCs at comparable levels and significantly enhanced senescent MSCs' immunosuppressive effect on T-cell proliferation. Our results define immune functional defects acquired by senescent MSCs, which are reversible by IFNγ prelicensing.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
References
-
- Phinney DG, Galipeau J, Krampera M, Martin I, Shi Y, Sensebe L. MSCs: science and trials. Nat Med. 2013;19(7):812. - PubMed
-
- Kallekleiv M, Larun L, Bruserud Ø, Hatfield KJ. Co-transplantation of multipotent mesenchymal stromal cells in allogeneic hematopoietic stem cell transplantation: a systematic review and meta-analysis. Cytotherapy. 2016;18(2):172-185. - PubMed
-
- Galipeau J. The mesenchymal stromal cells dilemma--does a negative phase III trial of random donor mesenchymal stromal cells in steroid-resistant graft-versus-host disease represent a death knell or a bump in the road? Cytotherapy. 2013;15(1):2-8. - PubMed
-
- Pittenger MF, Mackay AM, Beck SC, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143-147. - PubMed
-
- von Bahr L, Sundberg B, Lönnies L, et al. Long-term complications, immunologic effects, and role of passage for outcome in mesenchymal stromal cell therapy. Biol Blood Marrow Transplant. 2012;18(4):557-564. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
